











Севастополь, Россия
Севастополь, Россия
УДК 581.526.323 Бентос. Фитобентос
УДК 262.5 Вселенские соборы
Впервые приведены сведения о межгодовой динамике количественных показателей макрофитобентоса и проведен сравнительный анализ пространственно-временных изменений вклада доминирующих видов макрофитов в западной части Севастопольской бухты за 40-летний период. Гидроботанические исследования выполняли в летний период 1977, 2008 и 2017 гг. на одних и тех же разрезах по стандартной методике. Выявлено, что на протяжении изучаемого периода в составе донной расти-тельности сформировались полидоминантные фитосообщества, где господствуют виды, обитающие в среде с повышенным уровнем эвтрофирования. При этом отмечены высокая доля эпифитирующих водорослей и незначительная роль Gongolaria barbata. За исследуемый промежуток времени произошло поднятие нижней границы произрастания макрофитов и зарегистрировано резкое снижение биомассы макро-фитобентоса на глубине свыше 5 м. Установлено, что изменения в распределении и составе донной растительности в западной части бухты обусловлены ее геоэкологическим состоянием, которое зависит от воздействия антропогенных и природных факторов. Строительство в бухте гидротехнических сооружений приводит к изменению распространения зарослей морских трав, обитающих на мягких грунтах, и во-дорослей, встречающихся на твердом субстрате. Выявлено, что после экстремальных штормов в составе растительного покрова преобладают сезонные и однолетние виды водорослей, при этом на глубине 0.5–1 м отмечаются лишь ювенильные слоевища Gongolaria barbata. Полученные результаты могут быть использованы для мониторинговых исследований экологической ситуации в бухте, а также при организации прибрежно-морского природопользования.
прибрежная зона, донная растительность, морские травы, Gongolaria barbata, Чер-ное море, Севастополь, Севастопольская бухта
Введение
За последние годы возрастание объемов неочищенных стоков, неконтролируемая рекреационная нагрузка и активное строительство на побережье привели к увеличению уровня эвтрофирования прибрежной зоны Крымского полуострова [1–3]. Наиболее интенсивно антропогенное воздействие на экологическое состояние прибрежья проявляется в бухтах и портовых акваториях. Севастопольская бухта относится к акваториям активного хозяйственного использования. В настоящее время она выполняет роль резервуара, в которую поступают промышленные и хозяйственно-бытовые стоки, а также ливневые воды с площади водосбора. Ежедневно в бухту сбрасывается до 10–15 тыс. м3 неочищенных или условно-чистых вод, с которыми в нее попадает широкий спектр химических соединений органического и неорганического происхождения [4, 5]. Неблагоприятная экологическая ситуация в бухте вызывает изменения в составе и биомассе макрофитобентоса вплоть до его полной деградации в некоторых частях ее акватории.
Сведения о современном состоянии макрофитобентоса в Севастопольской бухте малочисленны [6, 7]. Впервые произрастание некоторых видов макрофитов в Черном море, в частности в Севастопольской бухте, было зарегистрировано в трудах Н. Н. Воронихина в начале XX в. Автор отметил, что водоросли и высшая водная растительность произрастают на определенном типе субстрата. В этот же период С. М. Переяславцева составила карту бухты, в которой указано распространение донных сообществ . Позднее С. А. Зернов представил карту распределения донной растительности в Севастопольской бухте . Автор показал, что в начале ХХ в. донные биоценозы бухты соответствовали общей схеме биоценозов Черного моря, где распределение макрофитобентоса в основном определялось субстратом (на твердых грунтах встречалась цистозира, а на мягких – морские травы) 4). В работе высказано предположение, что в бухте под влиянием антропогенной деятельности будут происходить существенные изменения состояния биоценозов 4). Действительно, уже в 1930-е гг. отмечено значительное загрязнение бухты, в составе местной фауны которой были выявлены заметные негативные изменения [8].
Впервые гидроботаническую съемку макрофитобентоса в Севастопольской бухте провела А. А. Калугина-Гутник в 1967 г. В материалах работы представлен видовой состав и рассчитана биомасса макрофитов по глубинам в разных частях бухты. Позднее, в 1977 г., А. А. Калугина-Гутник продолжила исследования донной растительности бухт Севастопольского взморья [9].
Показательно, что за последние 40 лет в связи с социально-экономи-
ческими причинами степень загрязнения бухты неоднократно менялась.
Поэтому возникает необходимость организации мониторинговых наблюдений за изменением состава и структуры макрофитобентоса, считающегося биоиндикатором состояния морской среды.
Цель работы – выявить особенности межгодовой динамики распределения макрофитобентоса в западной части Севастопольской бухты за 1977–2017 гг.
Материалы и методы исследования
Протяженность Севастопольской бухты составляет около 7.5 км при максимальной ширине 1 км (рис. 1). При входе в бухту глубина достигает 20 м, в вершине – 5 м. Бухта образовалась вследствие затопления устьевой части р. Черной в ходе послеледникового подъема уровня моря. Берега бухты высокие, сложены сарматскими известняками, береговая линия сильно изрезана. Прибрежный рельеф расчленен балками, продолжениями которых являются более мелкие бухты и вогнутости береговой линии. Тип берега – абразионно-бухтовый ингрессионный риасовый [10]. В современных условиях береговая зона существенно преобразована (бетонирование береговой линии, строительство молов, пирсов). Известно, что в 2022 г. непреобразованные берега бухты составляли лишь 1.1 км (3 % от первоначальной длины) береговой линии [11].
В настоящее время Севастопольская бухта представляет собой полузамкнутую акваторию эстуарного типа с затрудненным водообменом [5]. Вследствие своей конфигурации бухта подвержена воздействию волнения только западного направления. После строительства входных молов в конце 1970-х гг. значительное волнение в бухту почти не проникает [10]. Основными природными факторами, определяющими гидрохимическую структуру вод в бухте, являются взаимодействие с атмосферой, сток пресных вод р. Черной в восточную часть и приток соленых морских вод через входной пролив в ее западной части [5].
Гидроботанические исследования в Севастопольской бухте выполняли с применением легководолазного снаряжения и использованием маломерных судов в июле 2017 г. Распределение и биомассу макрофитов оценивали для прибрежной зоны западной части бухты (разрезы 1–4). (рис. 1). Координаты разрезов определяли при помощи портативного GPS-приемника (Oregon 650) (табл. 1).
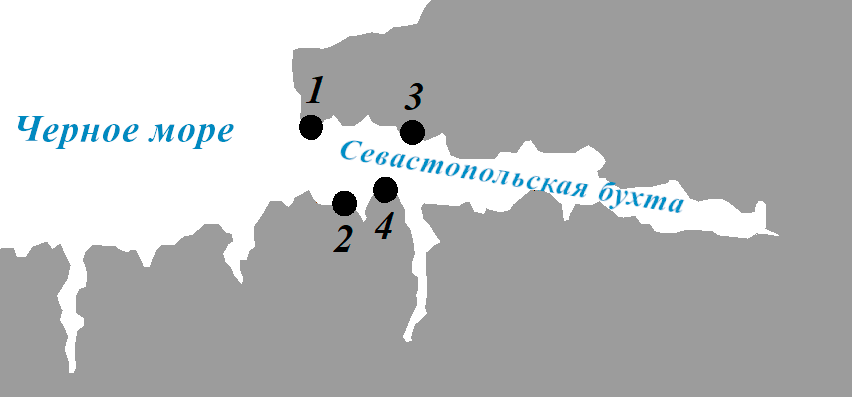
Рис. 1. Картосхема гидроботанических разрезов в западной части Севастопольской бухты (1 – мыс Конс-
тантиновский; 2 – мыс Хрустальный; 3 – мыс Славы;
4 – памятник затопленным кораблям)
Fig. 1. Schematic map of the location of hydrobota-
nical profiles in the western part of Sevastopol Bay (1 – Cape Konstantinovsky; 2 – Cape Khrustalny; 3 – Cape Slavy; 4 – Monument to Sunken Ships)
Таблица 1. Координаты разрезов, диапазон глубин, количество отобранных проб макрофитобентоса в Севастопольской бухте
Table 1. Coordinates and depth range of hydrobotanical profiles, number of sampled macrophytobenthos in Sevastopol Bay
|
Разрез / |
Координаты / |
Глубины, м / |
Количество проб / |
|||||
|
северная |
восточная |
0.5 |
1 |
3 |
5 |
7–10 |
||
|
1 |
44°37'36" |
33°30'44" |
+ |
+ |
+ |
+ |
– |
16 |
|
2 |
44°37'1" |
33°31'2" |
+ |
+ |
+ |
+ |
+ |
20 |
|
3 |
44°37'35" |
33°31'59" |
+ |
+ |
+ |
+ |
– |
16 |
|
4 |
44°37'3" |
33°31'29" |
+ |
+ |
+ |
+ |
– |
16 |
Примечание: тире – донная растительность отсутствует.
Note: dash – no bottom vegetation.
Для изучения состава макрофитобентоса отбирали пробы по стан-
дартной методике . На глубинах 0.5, 1, 3, 5, 10 и 15 м закладывали по четыре учетные площадки размером 25 × 25 см, отобрано 68 количественных проб (табл. 1). Донную растительность описывали согласно доминантной классификации по работе . При анализе структуры фитосообществ использовали индекс видового разнообразия Шеннона (Н). Водоросли идентифицировали по определителю с учетом последних номенклатурных изменений (URL: http://www.algaebase.org). Сведения о составе и распространении макрофитобентоса в бухте за 1977 и 2008 гг. получены одним из авторов, участвовавшим в сборе и обработке материала, выполненных в летний период в тех же районах по аналогичной методике.
Результаты и обсуждение
На основе проведенных исследований дана характеристика распределения макрофитобентоса и входящих в его состав доминирующих видов макрофитов в западной части Севастопольской бухты.
Распределение донной растительности в бухте в 1977 г. В устьевой части Севастопольской бухты на северном прибрежье на разрезе 1 (м. Константиновский) донная растительность зарегистрирована до глубины 5 м.
Максимальная общая биомасса макрофитов отмечена на глубине 3 м, на глубине 0.5 и 1 м количественный показатель был несколько ниже, тогда как на глубине 5 м биомасса снижалась в шесть раз (табл. 2). В изучаемом диапазоне глубин преобладала Gongolaria barbata (Stackhouse) Kuntze (= Cystoseira barbata), ее доля была высокой (рис. 2). В составе макрофитобентоса встречались Cladophora albida (Nees) Kütz. и Ulva rigida L. Эпифитирующие водоросли не обнаружены. Значения индекса видового разнообразия по глубинам колебались в широком диапазоне от 0.07 до 1.48. Низкие его значения на глубине 1 и 3 м объясняются тем, что на этих глубинах зафиксированы почти чистые заросли Gongolaria barbata (табл. 2).
Таблица 2. Изменение биомассы макрофитобентоса, доли входящих в его состав доминирующих и эпифитирующих видов макрофитов, индекса видового разнообразия (Н) в Севастопольской бухте по глубинам и годам
Table 2.Changes in the total biomass of macrophytobenthos, percentage of its dominant and epiphytic macrophyte species, diversity index (H) in Sevastopol Bay by depth and years
|
Разрез / Profile |
Глубина, м / |
Год / Уear |
Общая биомасса макрофитов, г·м–2 / Total biomass of macrophytes, g·m–2 |
Доля, % / |
Н |
|
|
Gongolaria barbata |
Эпифитов / |
|||||
|
1 |
0.5 |
1977 |
1608.4 ± 422.3 |
88 |
0 |
1.42 |
|
2008 |
1088.9 ± 251.5 |
0 |
5 |
2.19 |
||
|
2017 |
992.2 ± 302.1 |
63 |
3 |
1.58 |
||
|
1 |
1977 |
1382.6 ± 214.9 |
95 |
0 |
0.31 |
|
|
2008 |
1425.4 ± 396.4 |
0 |
11 |
1.99 |
||
|
2017 |
422.6 ± 58.1 |
73 |
15 |
1.41 |
||
|
3 |
1977 |
2249.8 ± 92.5 |
99 |
0 |
0.07 |
|
|
2008 |
361.6 ± 92.6 |
0 |
5 |
2.52 |
||
|
2017 |
424.3 ± 31.9 |
44 |
27 |
2.19 |
||
|
5 |
1977 |
370.1 ± 117.7 |
84 |
0 |
1.48 |
|
|
2008 |
296.5 ± 58.8 |
0 |
6 |
2.10 |
||
|
2017 |
18.6±7.7 |
18 |
26 |
2.39 |
||
|
7 |
2008 |
63.1 ± 18.7 |
20 |
23 |
3.37 |
|
|
2 |
0.5 |
1977 |
38.8 ± 5.2 |
0 |
0 |
0.64 |
|
2008 |
360.2 ± 40.4 |
0 |
4 |
2.03 |
||
|
2017 |
744.7 ± 330.3 |
70 |
9 |
1.76 |
||
|
1 |
1977 |
375.0 ± 128.9 |
0 |
0 |
0 |
|
|
2008 |
120.5 ± 60.2 |
3 |
6 |
2.54 |
||
|
2017 |
1224.3 ± 135.5 |
80 |
17 |
1.24 |
||
Продолжение таблицы
|
Разрез / Profile |
Глубина, м / |
Год / Уear |
Общая биомасса макрофитов, г·м–2 / Total biomass of macrophytes, g·m–2 |
Доля, % / |
Н |
|
|
Gongolaria barbata |
Эпифитов / |
|||||
|
2 |
3 |
1977 |
623.8 ± 45.1 |
0 |
0 |
0 |
|
2008 |
537.1 ± 194.4 |
39 |
2 |
2.31 |
||
|
2017 |
1310.2 ± 431.9 |
45 |
48 |
1.86 |
||
|
5 |
1977 |
600.9 ± 145.3 |
0 |
0 |
0.65 |
|
|
2008 |
399.3 ± 141.3 |
79 |
14 |
1.29 |
||
|
2017 |
306.3 ± 67.3 |
55 |
25 |
1.85 |
||
|
10 |
2008 |
50.0 ± 2.9 |
38 |
1 |
2.32 |
|
|
7 |
2017 |
13.1 ± 6.4 |
72 |
0 |
1.57 |
|
|
3 |
0.5 |
1977 |
623.3 ± 97.3 |
0 |
0 |
0.15 |
|
2008 |
789.9 ± 343.9 |
55 |
3 |
2.11 |
||
|
2017 |
4699.6 ± 1206.9 |
86 |
12 |
0.84 |
||
|
1 |
1977 |
588.5 ± 128.2 |
0 |
0 |
0 |
|
|
2008 |
253.3 ± 112.8 |
0 |
1 |
1.03 |
||
|
2017 |
5063.6 ± 346.6 |
87 |
7 |
0.88 |
||
|
3 |
1977 |
689.9 ± 130.7 |
0 |
0 |
0 |
|
|
2008 |
202.7 ± 75.9 |
0 |
2 |
0.49 |
||
|
2017 |
2322.8 ± 363.1 |
79 |
4 |
1.06 |
||
|
5 |
1977 |
200.7 ± 76.9 |
0 |
0 |
0.97 |
|
|
2008 |
228.4 ± 33.3 |
0 |
0 |
0.05 |
||
|
2017 |
1.2 ± 0.54 |
0 |
0 |
1.23 |
||
|
4 |
0.5 |
1977 |
917.0 ± 122.4 |
0 |
0 |
0.77 |
|
2008 |
955.9 ± 284.7 |
52 |
6 |
2.59 |
||
|
2017 |
5483.0 ± 1536.9 |
87 |
8 |
0.97 |
||
|
1 |
1977 |
1602.6 ± 127.4 |
0 |
0 |
0.02 |
|
|
2008 |
1406.2 ± 431.1 |
55 |
22 |
2.26 |
||
|
2017 |
3416.2 ± 1039.5 |
85 |
6 |
0.89 |
||
|
3 |
1977 |
1079.0 ± 543,7 |
0 |
0 |
0.82 |
|
|
2008 |
616.1 ± 172.1 |
50 |
4 |
2.06 |
||
|
2017 |
1863.7 ± 327.0 |
39 |
10 |
1.91 |
||
|
5 |
2008 |
270.9 ± 64.0 |
76 |
3 |
1.27 |
|
|
2017 |
655.2 ± 135.5 |
32 |
39 |
2.42 |
||
В устьевой части Севастопольской бухты на южном прибрежье на разрезе 2 (м. Хрустальный) на глубине 0.5–3 м доминировала Ulva rigida (рис. 2). Общая биомасса макрофитов в этом диапазоне при увеличении глубины возрастала в 16 раз (табл. 2). На глубине 5 м преобладала Zostera noltei Hornem.
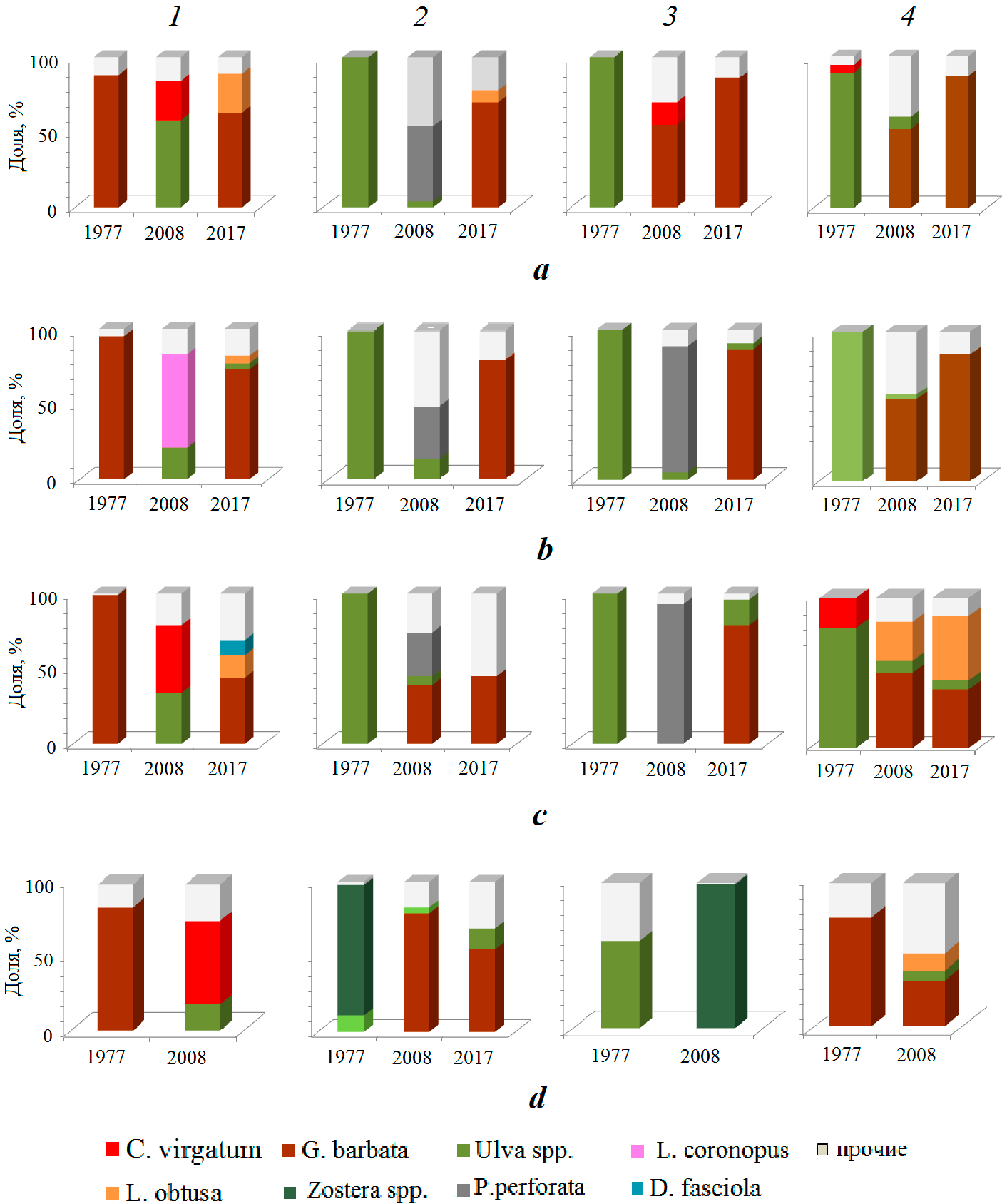
Рис. 2. Изменение доли доминирующих видов макрофитов по разрезам и годам на глубине 0.5 м (a), 1 м (b), 3 м (c) и 5 м (d) (1 – мыс Константиновский;
2 – мыс Хрустальный; 3 – мыс Славы; 4 – памятник затопленным кораблям)
Fig. 2. Change in the proportion of dominant macrophyte species by рrofile and years at the depth of 0.5 m (a), 1 m (b), 3 m (c) and 5 m (d) (1 – Cape Konstantinovsky; 2 – Cape Khrustalny; 3 – Cape Slavy; 4 – Monument to the Sunken Ships)
Среди зарослей морской травы отмечена Ulva rigida. Эпифиты отсутствовали. Значения индекса видового разнообразия были невысокими (0–0.65), что свидетельствует о монодоминантной структуре фитосообщества.
По мере продвижения вглубь бухты вдоль северного прибрежья на разрезе 3 (м. Славы) на глубине 0.5–5 м доминировала Ulva rigida (рис. 2). Общая биомасса макрофитов на глубине 0.5–3 м слабо варьировала, на глубине 5 м она снижалась примерно втрое (табл. 2). Показательно, что на глубине 1–3 м макрофитобентос представлен скоплениями Ulva rigida, тогда как на глубине 0.5 м в его составе отмечена Ulva sp., а на глубине 5 м – Bryopsis hypnoides Lamour. Значения индекса видового разнообразия свидетельствуют о преобладающем доминировании вида в альгосообществе.
Далее вглубь бухты вдоль южного прибрежья на разрезе 4 (памятник затопленным кораблям) на глубине 0.5–3 м также доминировала Ulva rigida (рис. 2). Максимальная общая биомасса макрофитов зарегистрирована на глубине 1 м, при этом на глубине 0.5 и 3 м этот показатель ниже в 1.7–1.5 раза. В составе донной растительности на всех глубинах встречались представители рода Ceramium. Значения индекса видового разнообразия варьировали по глубинам от 0.03 до 0.82.
Распределение донной растительности в бухте в 2008 г. На разрезе 1 на глубине 0.5 м преобладали Ulva intestinalis L., U. rigida и Ceramium virgatum Roth (рис. 2). В составе донной растительности также встречались Callithamnion corymbosum (Smith) Lyngb., Ceramium diaphanum (Lightf.) Roth. На глубине 1 м доминировали Laurencia coronopus J. Ag. и Ulva intestinalis. На этой глубине также отмечены Ulva rigida, Ceramium virgatum и виды рода Cladophora. В диапазоне глубин 3–5 м господствовали Ceramium virgatum и Ulva rigida. Доля этих видов изменялась в пределах 45–57 и 18–20 % общей биомассы макрофитов соответственно. Общая биомасса макрофитов на этих глубинах варьировала в нешироком интервале (табл. 2). Характерно, что при увеличении глубины от 0.5 до 5 м вклад Ulva intestinalis снижался с 35 до 1 % общей биомассы макрофитов. На глубине 7 м значение общей биомассы макрофитов было невысоким (табл. 2). Здесь доминировали Gongolaria barbata и Ulva rigida. На этой глубине обильно представлены эпифитирующие водоросли. Среди них преобладали Ceramium virgatum, Ectocarpus confervoides (Roth) Le Jolis, Laurencia coronopus и Vertebrata subulifera (C. Ag.) Kütz. Значения индекса видового разнообразия свидетельствуют о сложной полидоминантной структуре сообществ, в которых отмечен высокий вклад сопутствующих видов и водорослей-эпифитов.
На разрезе 2 на глубине 0.5–1 м доминировала Palisada perforata (Bory) K.W. Nam, ее доля с увеличением глубины в этом диапазоне снижалась с 50 до 36 % общей биомассы макрофитов (рис. 2), которая также уменьшалась в три раза (табл. 2). В составе донной растительности отмечены Gelidium crinale (Hare ex Turner) Gaillon, Ulva intestinalis, U. rigida, Callithamnion corymbosum. Представители рода Cladophora встречены как в литофитной, так и в эпифитной форме. На глубине 3 м преобладали Gongolaria barbata и Palisada perforata. В составе макрофитобентоса также обильно представлены Cladophoropsis membranacea (Ag.) Börg., Gelidium crinale, Ulva rigida.
На глубине 5–10 м доминировала Gongolaria barbata, доля которой с увеличением глубины снижалась вдвое, а общая биомасса макрофитов – в восемь раз (табл. 2). Под пологом и среди слоевищ Gongolaria barbata на глубине 5 м отмечена Ulva rigida, а на глубине 10 м – Cladostephus spongiosus (Huds.) C. Ag., Laurencia coronopus и Cladophora laetevirens (Dillw.) Kütz. В составе донной растительности на глубине 5–10 м встречены глубоководные виды (Zanardinia typus (Nardo) P. C. Silva, Carradoriella elongata (Huds.) Savoie & G.W. Saunders., Nereia filiformis (J. Ag.) Zanard.). Наибольшее развитие эпифитная синузия получила на глубине 5 м. Среди эпифитных водорослей преобладали Vertebrata subulifera и Stilophora tenella (Esper) P.C. Silva. Значения индекса видового разнообразия свидетельствуют о высокой доле сопутствующих видов в структуре фитосообщества.
На разрезе 3 на глубине 0.5 м преобладала Gongolaria barbata (рис. 2). В составе макрофитобентоса обильно представлены Ceramium virgatum, Carradoriella denudata (Dillw.) Savoie et G. W. Saunders, виды родов Cladophora и Ulva. На глубине 1–3 м доминировала Palisada perforata, на долю этого вида приходилось 84–93 % общей биомассы макрофитов. С увеличением глубины вклад Cladophora laetevirens возрастал с 1 до 5 %, а Ulva rigida – снижался с 5 до 1 % общей биомассы макрофитов. На глубине 5 м доминировала Zostera marina. Вклад эпифитирующих водорослей на всех исследованных глубинах был незначительный (табл. 2). Значения индекса видового разнообразия варьировали в широком интервале и зависели от сложности структуры сообщества.
На разрезе 4 на глубине 0.5–5 м преобладала Gongolaria barbata (рис. 2). Максимальная общая биомасса макрофитов отмечена на глубине 1 м, тогда как на глубине 0.5 м этот показатель был ниже в 1.5 раза, а глубже 1 м уменьшался в два и пять раз на глубине 3 и 5 м соответственно (табл. 2). В составе донной растительности на всех исследованных глубинах встречены Ulva rigida, Cladophoropsis membranacea. На глубине 0.5–1 м была велика доля G. crinale и Dermocorynus dichotomus (J. Ag.) Gargiulo Morabito and Manghisi, на глубине 3 м – литофитной формы Laurencia obtusa (Huds.) J. V. Lamour., а на глубине 5 м – Zanardinia typus. Вклад Cladophora laetevirens при увеличении глубины в изучаемом диапазоне снижался с 9 до 1 %. Наиболее обильно эпифиты представлены на глубине 1 м. Среди них доминировали Vertebrata subulifera и виды рода Cladophora. Высокие значения индекса видового разнообразия свидетельствуют о полидоминантной структуре сообщества.
Распределение донной растительности в бухте в 2017 г. В устьевой части Севастопольской бухты на разрезе 1 донная растительность зафиксирована до глубины 5 м. На глубине 0.5–3 м преобладала Gongolaria barbata (табл. 2). Доля этого вида была максимальной на глубине 1 м, тогда как по мере увеличения глубины этот показатель снижался в два и четыре раза. Общая биомасса макрофитов при повышении глубины уменьшалась более чем в 50 раз. В составе макрофитобентоса на всех исследуемых глубинах встречена Dictyota fasciola (Roth) Howe (рис. 2). Доля этого вида на глубине 0.5–1 м варьировала в пределах 2–4 %, тогда как на глубине 3–5 м она резко возрастала и составляла 10–28 % общей биомассы макрофитов. Среди зарослей Gongolaria barbata на глубине 0.5 м отмечена литофитная форма Laurencia obtusa, глубже этот вид встречался как эпифит. На глубине 5 м обильно представлена Cladophora serisea (Huds.) Kütz. Вклад эпифитирующих водорослей существенно возрастал с увеличением глубины. Изменение индекса видового разнообразия свидетельствует об усложнении структуры сообщества при увеличении глубины.
На разрезе 2 на глубине 0.5–5 м доминировала Gongolaria barbata (рис. 2). Общая биомасса макрофитов возрастала почти в два раза с увеличением глубины от 0.5 до 3 м (табл. 2). При дальнейшем повышении глубины этот показатель снижался более чем в четыре раза, а на глубине 7 м донная растительность практически отсутствовала. Доля доминирующего вида на глубине
0.5–1 м изменялась от 70 до 80 %, тогда как на глубине 3–5 м – уменьшалась до 45–55 % общей биомассы макрофитов. В составе макрофитобентоса на всех глубинах среди зарослей Gongolaria barbata встречены зеленые водоросли родов Cladophora и Ulva. На глубине 3–5 м отмечены глубоководные виды Nereia filiformis и Carradoriella elongata. Вклад эпифитов с увеличением глубины возрастал от 9 до 17 % (глубина 0.5 и 1 м), а с дальнейшим ее увеличением снижался от 48 до 25 % общей биомассы макрофитов (глубина 3 и 5 м). Среди эпифитирующих водорослей преобладала Vertebrata subulifera. Индекс видового разнообразия по глубинам изменялся от 1.24 до 1.86.
На разрезе 3 донная растительность обнаружена до глубины 3 м (табл. 2). На этих глубинах доминировала Gongolaria barbata (рис. 2). Общая биомасса макрофитов на глубине 0.5–1 м достигала высоких значений (от 4699.6 ± 1206.9 до 5063.6 ± 346.6 г·м–2), на глубине 3 м она снижалась в два раза, тогда как на глубине 5 м макрофитобентос практически отсутствовал. В составе макрофитобентоса с увеличением глубины возрастал вклад Ulva rigida от 1 до 17 %, тогда как роль эпифитирующих водорослей уменьшалась с 12 до 4 % общей биомассы макрофитов. Среди эпифитов преобладала Shacelaria cirrhosa (Roth) C. Ag. На глубине 5 м единично встречались Dictyota fasciola, Ulva rigida и Carradoriella elongata. Индекс видового разнообразия на глубине 0.5–3 м варьировал в нешироком интервале (0.84–1.06), что свидетельствует о преобладании доминирующего вида.
На разрезе 4 на глубине 0.5–5 м преобладала Gongolaria barbata. Общая биомасса макрофитов с увеличением глубины в изучаемом диапазоне снижалась в восемь раз, а доля доминирующего вида уменьшалась почти втрое (табл. 2). В составе донной растительности на всех глубинах отмечены Cladophora laetevirens и Ulva rigida. Ульва на глубине 0.5–1 м встречалась единично, а на глубине 3–5 м ее доля возрастала до 6–7 % общей биомассы макрофитов (рис. 2). На глубине 3 м обильно представлена литофитная форма Laurencia obtusa, тогда как на глубине 5 м этот вид эпифитировал на Gongolaria barbata. Максимальный вклад эпифитирующих водорослей зарегистрирован на глубине 5 м, на глубине 0.5–3 м этот показатель был существенно ниже. Среди эпифитов преобладала Vertebrata subulifera. Значения индекса видового разнообразия показывают, что при увеличении глубины происходит усложнение структуры сообщества.
В конце 1970-х гг. западная часть Севастопольской бухты представляла собой акваторию, подверженную значительному антропогенному воздействию. В этот период в исследуемой части бухты (разрезы 2–4) зафиксировано обильное развитие зеленых водорослей, среди которых преобладала Ulva rigida.
Характерно, что этот вид произрастает в воде с высоким уровнем загрязнения хозяйственно-бытовыми стоками. Вероятно, на видовой состав макрофитобентоса повлиял выпуск канализации с городской застройки северной стороны, действующий с 1964 г. Кроме этого, в районе Северной бухты располагался рыбколхоз, деятельность которого также негативно влияла на донные биоценозы. Так, в работе [8] показано, что в местах стоянки судов и кораблей количество органического и аммонийного азота в донных осадках в 1.5 и 5 раз больше, чем в открытом море.
В период строительства северной части мола на разрезе 1 макрофитобентос представлен в основном зарослями Gongolaria barbata (84–99 % общей биомассы макрофитов) – ценозообразующим видом прибрежной зоны Черного моря (табл. 2). Хотя в 1977 г. на этом участке еще происходил частичный водообмен с открытой частью моря, общая биомасса макрофитов по глубинам была в 3–10 раз ниже, чем в этом же году у открытого прибрежья б. Омега, где доля Cystoseira spр. также достигала 95–98 % [9].
За прошедшие более чем 30 лет (1977–2008 гг.) в изучаемом районе бухты существенно изменился состав доминирующих видов макрофитов. На разрезах 2–4, где в 1977 г. господствовали виды ульвы, в 2008 г. на некоторых глубинах стала преобладать Gongolaria barbata (табл. 2). Вероятно, такая смена доминирующих видов говорит о некотором снижении степени загрязнения воды. В пользу этого предположения могут служить исследования, которые изложены в монографии [12]. В работе показано, что в конце XX в. в бухте уровни концентрации фосфатов, биогенных элементов, свидетельствующих прежде всего о выпуске сточных и ливневых вод, были заметно ниже, чем в период 1974–1983 гг. Кроме того, отмечено, что в 1998–1999 гг. количество фосфатов было в 16 раз ниже, чем в 1970-х гг.
В монографии [8] указано, что содержание нефтяных углеводородов и хлороформного битумоида, являющихся стойкими органическими загрязнителями донных осадков, в 1979–1985 гг. было несколько выше, чем в 1997–2000 гг. (328–999 и 451–507 мг/100 г; 0.82–2.7 и 1.21–1.25 г/100 г). Установлено [13], что в центральной части Севастопольской бухты в толще донных осадков на глубине от 5 до 20 см сформировалась зона чрезвычайно высокой концентрации полихлорированных бифенилов (ПХБ), до 600 нг·г–1 (в пересчете на сухую массу). В поверхностном слое донных осадков этого района значение ПХБ было несколько ниже (от 200 до 450 нг·г–1), что привело авторов к выводу об уменьшении техногенного загрязнения бухты.
При отборе проб макрозообентоса в июне 2009 г. на станциях, расположенных по всей акватории бухты, было установлено, что качество среды в районе памятника затопленным кораблям (разрез 4) оценивалось как «хорошее» в соответствии с индексом M-AMBI, тогда как на других станциях (центральная и восточная часть бухты) оно характеризовалось как «умеренное» или «бедное» [14]. Так, по сведениям ряда исследователей, в конце 1990-х – начале 2000-х гг. в экологической обстановке Севастопольской бухты наблюдалось улучшение. Однако это произошло не в результате природоохранных действий, а в связи с уменьшением объемов стоков промышленных предприятий из-за прекращения их работы, а также из-за уменьшения нефтяного загрязнения вследствие сокращения военно-морского флота [8, 12].
Тем не менее на исследуемых участках, особенно на разрезе 1, стали повсеместно произрастать водоросли (Ulva intestinalis, U. rigida, Cladophora laetevirens, Cl. serisea Ceramium virgatum, C. diaphanum, Callithamnion corymbosum, Carradoriella denudata, Ectocarpus confervoides), которые обитают в воде с повышенным уровнем эвтрофирования, что свидетельствует о наличии загрязняющих веществ в бухте.
Характерно, что с возведением при входе в бухту северной (длина 250 м) и южной (длина 500 м) частей мола изменился вдольбереговой поток наносов [11]. Этим можно объяснить смену субстрата на некоторых участках западной части бухты. Согласно авторам, ранее аккумулятивные формы в виде песчаных отмелей имелись в Северной и Александровской бухтах. Сейчас в первой бухте поступление песка полностью прекратилось, а во второй – сохранились лишь фрагменты отмелей [11]. Вероятно, из-за отсутствия мягких грунтов исчезли заросли морской травы (Zostera noltei) в районе м. Хрустального (разрез 2), которые ранее (1977 г.) были отмечены на глубине 5 м. В 2008 г. на этом разрезе на этой глубине обнаружены виды водорослей, произрастающие исключительно на твердом субстрате. В то же время появились незначительные скопления Zostera marina на глубине 5 м в районе м. Славы (разрез 3).
Возможно, что в 2008 г. на распространение и состав макрофитобентоса в западной части бухты оказали влияние последствия шторма. Известно, что в результате активной штормовой деятельности происходят деградация и разрушение прибрежных биоценозов. Так, в районе Карадага после сильнейшего шторма, зафиксированного в 1992 г., на глубине 0–10 м отмечено полное уничтожение донной растительности [15]. В ноябре 2007 г. в акватории Черного моря был зарегистрирован экстремальный шторм, при котором сила ветра составляла 27–32 м·с–1, а высота волн достигала 4 м [16]. Ранее было показано негативное воздействие этого шторма на состояние макрофитобентоса в Ласпинской бухте [17]. Общеизвестно, что наиболее интенсивный рост макрофитов в Черном море начинается только в весенне-летний период 8). Вероятно, поэтому донная растительность в прибрежной зоне этой части Севастопольской бухты летом 2008 г. характеризовалась высокой видовой мозаичностью, в составе макрофитобентоса массово встречались однолетние виды водорослей (Ceramium spp., Cladophora spp.) В этот период на изучаемых участках бухты обильно представлены водоросли рода Laurencia (L. coronopus, L. obtusa, L. papillosa = Palisada perforata) (рис. 2). Известно, что активный рост этих видов начинается при интенсивном солнечном освещении и с началом прогрева водной толщи [18]. Возможно, на месте уничтоженных штормом зарослей Gongolaria barbata, талломы которой растут довольно медленно, первоначально появились слоевища представителей рода Laurencia 8) [18]. Так, в акватории разреза 2 (м. Хрустальный) на глубине 0.5–1 м зарегистрированы лишь проростки Gongolaria barbata и только на глубине 3–5 м доля этого вида составляла 39–79 % общей биомассы макрофитов (табл. 2).
Проведенные в 2017 г. исследования выявили, что для западной части Севастопольской бухты характерна тенденция к снижению количественных показателей макрофитобентоса в нижней сублиторальной зоне, наблюдаемая в прибрежье Черного моря в районах с повышенным уровнем эвтрофирования [19, 20]. В бухтах эта тенденция приобретает катастрофические последствия. Так, в районе м. Константиновского (разрез 1) на глубине 5 м общая биомасса макрофитов уменьшилась в 20 раз, а доля Gongolaria barbata – в 5 раз с 1977 по 2017 г. (табл. 2). За этот же промежуток времени в районе м. Славы (разрез 3) общая биомасса макрофитов на этой же глубине снизилась более чем на два порядка. На остальных исследуемых участках также отмечено уменьшение вклада Gongolaria barbata за период с 2008 по 2017 г.
Характерно, что в настоящее время на всех разрезах западной части бухты в составе макрофитобентоса обильно представлены эпифитирующие водоросли (Vertebrata subulifera, виды рода Cladophora), обладающие высокой конкурентной способностью, которая определяется их устойчивостью к изменяющимся условиям среды, быстрым ростом, способностью эффективно ассимилировать избыточное количество органических и минеральных элементов .
Таким образом, в западной части Севастопольской бухты происходящие изменения в составе донной растительности, вероятно, обусловлены воздействием как природных факторов, так и хозяйственной деятельностью:
– геоэкологическая ситуация в бухте, связанная с высокой антропогенной нагрузкой, привела к тому, что в составе макрофитобентоса доминирующая роль стала принадлежать видам, произрастающим в среде с повышенным уровнем загрязняющих веществ, при этом отмечена высокая доля эпифитирующих водорослей, тогда как вклад Gongolaria barbata снижается. Кроме этого, наблюдается резкое уменьшение количественных показателей растительной компоненты на глубине свыше 5 м. Значения индекса видового разнообразия свидетельствуют о сложной полидоминантной структуре фитосообществ;
– строительство в бухте гидротехнических сооружений, которые изменили вдольбереговой поток наносов, привело к перераспределению распространения зарослей морских трав, обитающих на мягких грунтах, и водорослей, встречающихся на твердом субстрате;
– увеличение интенсивности штормов негативно влияет на состояние донных сообществ. Выявлено, что после экстремальных штормов растительный покров отличается значительной видовой мозаичностью, преобладанием сезонных и однолетних видов водорослей, при этом на глубине 0.5–1 м отмечены лишь ювенильные слоевища Gongolaria barbata.
Заключение
Показано распределение биомассы макрофитобентоса и входящих в его состав доминирующих видов макрофитов по глубинам и годам (1977, 2008 и 2017 гг.) в западной части Севастопольской бухты.
Проведен сравнительный анализ пространственно-временных изменений вклада преобладающих видов макрофитов за 40-летний период.
Выявлено, что в составе донной растительности на протяжении исследуемого периода сформировались полидоминантные фитосообщества, где доминируют виды, обитающие в среде с повышенным уровнем эвтрофирования, при этом отмечены высокая доля эпифитирующих водорослей и незначительная роль Gongolaria barbata. Характерно резкое снижение количественных показателей макрофитобентоса на глубине свыше 5 м, наблюдается поднятие нижней границы произрастания макрофитов.
Установлено, что изменения в распределении и составе донной растительности в западной части Севастопольской бухты обусловлены ее геоэкологическим состоянием, которое зависит от воздействия антропогенных и природных факторов.
Полученные результаты могут быть использованы для мониторинговых исследований экологической ситуации в бухте, а также при организации прибрежного и морского природопользования.
1. Основные источники загрязнения морской среды Севастопольского региона / Е. И. Овсяный [и др.] // Экологическая безопасность прибрежной и шельфовой зон и комплексное использование ресурсов шельфа. Севастополь : ЭКОСИ-Гидрофизика, 2001. Вып. 2. С. 138–152. EDN KQOLRV.
2. Источники загрязнения прибрежных вод Севастопольского района / В. М. Гру-зинов [и др.] // Океанология. 2019. Т. 59, № 4. С. 579–590. EDN VEOVBB. https://doi.org/10.31857/S0030-1574594579-590.
3. Вержевская Л. В., Миньковская Р. Я. Структура и динамика антропогенной нагрузки на прибрежную зону Севастопольского региона // Экологическая безопасность прибрежной и шельфовой зон моря. 2020. № 2. С. 92–106. EDN XGZGZL. https://doi.org/10.22449/2413-5577-2020-2-92-106
4. Слепчук К. А., Совга Е. Е. Уровень эвтрофирования восточного района Севасто-польской бухты // Экологическая безопасность прибрежной и шельфовой зон моря. 2018. № 2. С. 53–59. EDN YLLQLJ. https://doi.org/10.22449/2413-5577-2018-2-53-59
5. Совга Е. Е., Мезенцева И. В., Хмара Т. В. Природно-климатические и антропоген-ные факторы, определяющие самоочистительную способность морских мелко-водных экосистем в отношении восстановленных форм азота // Экологическая безопасность прибрежной и шельфовой зон моря. 2021. № 3. С. 23–36. EDN YUWVPQ. https://doi.org /10.22449/2413-5577-2021-3-23-36
6. Евстигнеева И. К., Танковская И. Н. Современное состояние и изменчивость мак-рофитобентоса ботанического памятника природы «Ушакова балка» (Черное море, Севастопольский регион) // Современные проблемы экологии Азово-Черноморского региона : материалы IV Международной конференции. 8–9 ок-тября 2008, Керчь. Керчь : Изд-во ЮгНИРО, 2008. C. 92–98.
7. Евстигнеева И. К., Евстигнеев В. П., Танковская И. Н. Структурно-функциональные особенности черноморского макрофитобентоса в районах с разным ветро-волновым режимом // Вода и экология: проблемы и решения. 2019. № 2. С. 82–91. EDN UFYZRF. https://doi.org/10.23968/2305-3488.2019.24.2.82-91
8. Миронов О. Г., Кирюхина Л. Н., Алемов С. В. Санитарно-биологические аспекты экологии севастопольских бухт в ХХ веке. Севастополь : ЭКОСИ-Гидрофизика, 2003. 185 с. EDN XXFQVL.
9. Калугина-Гутник А. А. Изменения в донной растительности Севастопольской бухты за период с 1967 по 1977 г. // Экология моря. 1982. Вып. 9. С. 48–62. EDN ZBPSLT.
10. Горячкин Ю. Н., Долотов В. В. Морские берега Крыма. Севастополь : Колорит, 2019. 256 с. EDN ARVKTY.
11. Ефремова Т. В., Горячкин Ю. Н. Морфодинамика севастопольских бухт под воздействием антропогенной деятельности // Экологическая безопасность прибрежной и шельфовой зон моря. 2023. № 1. С. 31–47. EDN THAAMX. https://doi.org/10.29039/2413-5577-2023-1-31-47
12. Акватория и берега Севастополя: экосистемные процессы и услуги обществу. Севастополь : Аквавита, 1999. 290 с.
13. Малахова Л. В., Малахова Т. В., Егоров В. Н. Донные отложения морской аквато-рии и пресных водоемов Крыма как депо стойких органических загрязнителей // «Ломоносовские чтения–2019» : ежегодная научная конференция МГУ. Сева-стополь, 3–4 апреля 2019 г. Севастополь : Филиал МГУ в г. Севастополе, 2019. С. 209–210. EDN RYQMKJ.
14. Особенности пространственного распределения нефтяных углеводородов и структуры макрозообентоса бухт Севастопольская и Варна / Т. С. Осадчая [и др.] // Системы контроля окружающей среды. Севастополь, 2010. Вып. 13. С. 247–255. EDN ZTYQFF.
15. Костенко Н. С., Дикий Е. А., Заклецкий А. А. Тенденции многолетних изменений фитоценозов «цистозирового пояса» Карадагского природного заповедника (Крым, Черное море) // Морской экологический журнал. 2008. Т. 7, № 3. С. 25–36. EDN TYNTTZ.
16. Доценко С. Ф., Иванов В. А. Морские катастрофические природные явления Азо-во-Черноморского региона // Экологическая безопасность прибрежной и шель-фовой зон и комплексное использование ресурсов шельфа. Севастополь : ЭКО-СИ-Гидрофизика, 2011. Вып. 24. С. 209–218. EDN WDNWFT.
17. Pankeeva T. V., Mironova N. V. Spatiotemporal Changes in the Macrophytobenthos of Laspi Bay (Crimea, Black Sea) // Oceanology. 2019. Vol. 59, iss. 1. P. 86–98. EDN ZEOZVJ. https://doi.org/10.1134/S0001437019010168
18. Евстигнеева И. К. Сезонная динамика структуры ценопопуляций лауренций в Севастопольской бухте // Экология моря. 1983. Вып. 14. С. 56–62. EDN WIEZNF.
19. Мильчакова Н. А., Миронова Н. В., Рябогина В. Г. Морские растительные ресурсы // Промысловые биоресурсы Черного и Азовского морей. Севастополь : ЭКОСИ-Гидрофизика, 2011. Гл. 4. С. 117–139. EDN WPIJMN.
20. Миронова Н. В., Панкеева Т. В. Пространственно-временные изменения макрофи-тобентоса бухты Круглая (Черное море) // Юг России: экология, развитие. 2020. Т. 15, № 2. С. 125–139. EDN RWQGFF. https://doi.org/10.18470/1992-1098-2020-2-125-139