












Россия
Россия
Россия
Россия
Анализируются многолетние изменения экосистемы Черного моря, выражающиеся, кроме прочего, в потеплении вод на фоне антропогенного загрязнения и эвтрофикации. Цель работы заключается в описании структурных изменений водных масс, биотопов, а также в оценке изменений ключевых компонентов экосистемы. На основе анализа многолетних массивов данных о гидрологии, математического моделирования и гидробиологических исследований показано, что для полного понимания изменений экосистемы Черного моря необходимо учитывать не только потепление климата, антропогенное загрязнение и эвтрофикацию, но ихорические изменения структуры водных масс и связанных с ними биотопов. Продолжающаяся деоксигенация сокращает слой обитания холодноводных видов, а потепление уже привело к тому, что холодный промежуточный слой, с которым эти виды ассоциированы, исчез и растворился в окружающих водах при температуре около 9 °С. Дальнейшее потепление поверхностных вод может привести к деградации комплекса холодноводных видов, создающих трофическую основу современной экосистемы Черного моря. Сле- дует ожидать ее существенной трансформации вследствие увеличения роли тепло- водных и эвритермных видов
Черное море, экосистема, изменения климата, деоксигенация, водные массы, биотопы
Введение
Около 130 лет назад Н. И. Андрусов 1) впервые показал, что Черное море (ЧМ) представляет собой двухслойную систему, состоящую из тонкого (80– 200 м) «живого» кислородного слоя и сероводородной зоны. До конца XX в. считалось, что между этими двумя слоями имеется слой сосуществования кислородных и сероводородных вод или С-слой. Первые публикации об от- сутствии этого слоя [1] были восприняты очень осторожно. Однако исполь- зование новых методик в конце ХХ в. показало, что представления об С-слое были ошибочными [1–3]. Выяснилось, что между кислородной зоной и сероводородным слоем находится водная масса (30–50 м) трансформированной средиземноморской воды (O2 < 10 мкМ, H2S < 3 мкМ), или субкислородный слой (СКС) [1, 2]. Положение этого слоя в толще воды зависит от интенсив- ности вертикального водообмена и обилия растворенного органического ве- щества. Увеличение поступления питательных веществ в р. Дунай, Днепр и Дон в 10 раз с 1960-х по 1990-е гг. привело к антропогенной эвтрофикации с образованием большого количества опускающегося органического веще- ства [2]. Вертикальный водообмен не обеспечивал достаточной аэрации, чтобы компенсировать потребление кислорода для окисления этой массы органического вещества, и кислородный слой моря стал «обмелевать» [2, 4]. Глубина проникновения кислорода к верхней границе СКС изменилась со 130 м в 1955 г. до 90 м в 2013 г. [5]. К началу ХХ в. эвтрофикация моря уменьшилась [6], но из-за повышения температуры поверхностных вод деок- сигенация глубин продолжилась [7].
Таким образом, в последние годы в глубоководной зоне ЧМ произошли значительные изменения структуры биотопов, которые существенно повлия- ли на условия жизни планктона и рыб. Поэтому для полноценного понимания современных изменений экосистемы ЧМ необходимо учитывать не только такие факторы, как изменение климата, антропогенное загрязнение и эвтро- фикация, но и такие, как значительные изменения гидролого-гидрохимичес- ких и хорических свойств водных масс и биотопов. (Хорические изменения водных масс (от греч. chora – «место») наблюдаются в тех случаях, когда водная масса меняет свое пространственное положение и под воздействием другого сочетания внешних факторов в новом местоположении приобретает иные экологические свойства [8].)
Существующие гидролого-гидрохимические представления о структуре поверхностных водных масс в глубоководных районах ЧМ можно предста- вить следующим образом:
- Верхняя черноморская водная масса (ВЧВМ): T > 20 °С, S = 18.0…18.4 ЕПС, σt < 14 2), 3).
- Холодный промежуточный слой (ХПС): T < 8 °С, S = 18.0…19.0 ЕПС, σt = 14.0…14.8 2), 3).
- Субкислородный слой (СКС): T > 8 °С, σt = 15.8…16.2 [2, 3].
- Промежуточная черноморская водная масса (ПЧВМ): T > 8 °С, S > 20 ЕПС, σt = 16.2…17.9 2), 3). (Прибрежная и глубинная водные массы в данной работе не обсуждаются, поскольку речь идет только о поверхностных водах глубо- ководной зоны.)
Структуру биотопов в глубоководных районах ЧМ можно представить следующим образом:
- Биотоп поверхностной пленки (аэроконтур): сообщество аэробных ор- ганизмов поверхностной пленки – нейстон. Состав – постоянные или временные обитатели слоя 0–5 см: бактерии и простейшие, фитопланктон, зоопланк- тон, икра и личинки беспозвоночных и рыб [9, 10].
- Биотоп ВЧВМ (аэробная зона): сообщество аэробных организмов над се- зонным термоклином – эпибиота. (Термины «эпибиота» и «батибиота» мы использовали вслед за Т. С. Петипа 4), впервые описавшей эпи- и батипланк- тон.) Состав – теплолюбивые и эвритермные виды, населяющие верхнюю водную массу: фитопланктон, эпипланктон, макропланктон и основная масса рыб (Engraulis encrasicolus Linnaeus, 1758 и пр.) 4) [6, 11].
- Биотоп ХПС (аэробная зона): сообщество аэробных организмов под се- зонным термоклином – батибиота 4). Состав – батипланктон и холодноводные рыбы (Sprattus sprattus (Linnaeus, 1758) и др.), эвритермные виды планкто- на 4), 5) [11–14].
- Биотоп СКС (анаэроконтур): сообщество аэробных и анаэробных мик- роорганизмов субкислородного слоя. (Термин «анаэроконтур» впервые вве- ден В. В. Мельниковым [14].) Состав – аэробные и анаэробные виды цианобак- терий, денитрифицирующих бактерий, метанотрофы, метаногены, бактерии, восстанавливающие железо, бактерии, восстанавливающие марганец [14].
- Биотоп ПЧВМ (анаэробная зона): сообщество анаэробных микроорга- низмов сероводородной зоны. Состав – тионовые бактерии (до 40 % биомассы от всего бактериопланктона), пурпурные и ферментирующие бактерии, мета- нотрофы, метаногены, марганецредуцирующие бактерии, анаммоксибактерии и различные археи 6) [14]. Прибрежная и глубинная водные массы не обсужда- ются, поскольку речь идет только о поверхностных водах глубоководной зоны. В последние годы описанные выше представления о структуре водных масс и биотопов уже не соответствуют реальному положению дел из-за про- должительного потепления поверхностных вод 7). К настоящему времени в результате деоксигенации концентрация кислорода в глубинах моря сокра- тилась на 44 % [5]. Температура ХПС начала повышаться, и к 2019 г. эта вод- ная масса исчезла [15]. Информация об этом катастрофическом для экосисте- мы ЧМ событии, безусловно, требует неоднократной проверки на разных ря- дах данных. Дальнейший нагрев поверхностных вод может вызвать каскад- ные изменения всей экосистемы ЧМ, поскольку, например, холодноводные копеподы составляют основную часть кормового зоопланктона, основу раци- она мелких планктоноядных рыб ЧМ [12–16]. Кроме того, остается нерешен- ным один из основополагающих вопросов о том, как сокращение общей тол- щины кислородного слоя в глубоководных районах моря повлияло на хори
ческие изменения структуры биотопов пелагиали. Настоящее исследование в какой-то степени восполняет этот пробел.
Целью работы является анализ базовых процессов долгосрочных изме- нений экосистемы ЧМ под воздействием потепления, антропогенного за- грязнения, эвтрофикации и изменений структуры водных масс и оценка из- менений ключевых компонентов экосистемы.
.
Материалы и методы
Гидродинамический анализ был основан на модели океанской циркуля- ции NEMO версии 3.6 8). Эта модель была реализована в регионе ЧМ с разре- шением по горизонтали 0.037° × 0.028° и 31 неравномерно расположенным вертикальным уровнем. Онлайн-модель связана со схемой ассимиляции OceanVar [17, 18]. Наблюдения, включенные в BS-REA, содержат профили температуры/солености (T/S), полученные на месте с помощью SeaDataNet и CMEMS INS TAC, а также аномалии уровня моря. Обобщаются все доступ- ные данные наблюдений с судов (SYNOP SHIP), полученные с батитермогра- фов (BATHY), с дрейфующих буев (DRIBU), а также данные, извлеченные из оперативного архива ECMWF.
В этой статье мы рассмотрели пространственно-временны́ е колебания температуры ХПС и окружающих его вод за последние 30 лет. Долгопериод- ные и пространственные изменения температуры поверхности моря (ТПМ)
были рассчитаны на основе ежедневных спутниковых карт с разрешением 0.05° × 0.05°, охватывающих 40-летний период с 1982 по 2021 г. Вся инфор- мация была получена из службы мониторинга морской среды Copernicus (https://www.copernicus.eu) 9). Кроме того, мы использовали новые данные, полученные в ходе последних крупномасштабных экспедиций Центра кол- лективного пользования «НИС Профессор Водяницкий» Института биологии южных морей им. А. О. Ковалевского РАН в 2017–2023 гг. (с использованием CTD-зонда Idronaut Ocean Seven 320Plus M).
Для исследования многолетних изменений вертикального положения плотностных границ СКС были использованы материалы из западно- центральной части Черного моря. Распределение гидрохимических парамет- ров в глубоководной части моря носит квазипостоянный изопикнический ха- рактер [7], поэтому оценка многолетних изменений вертикального положения границ СКС проводилась в шкале условной плотности. Эти границы в сред- нем соответствуют изопикническим поверхностям: σt = 15.8 – верхняя грани- ца и σt = 16.2 – нижняя [2, 3]. Использование такого подхода решает две важ- ные методические проблемы:
- дефицит гидрохимических данных – данных по гидрохимии западной глубоководной зоны явно недостаточно для оценки их многолетних измене- ний (гидрологических станций почти на порядок больше);
- влияние случайных факторов на точность оценки – использование плотностных характеристик позволяет нивелировать действие случайных процессов и получать более достоверные количественные оценки эволюции экологической структуры вод ЧМ.
Западная часть моря была выбрана, поскольку сезонная динамика верти- кального положения изопикн в этом круговороте незначительна (около 5 м) по сравнению с восточным круговоротом, где она достигает 20 м. Если учесть, что межгодовая изменчивость также больше в восточном круго- вороте, то плотностные характеристики для определения долгосрочных изменений в западно-центральной части моря более статистически надежны.
Данные для расчетов были собраны в океанографических рейсах науч- ных судов СССР, России, Украины, Турции, Болгарии, Румынии. Для реана- лиза были использованы массивы информации из баз данных WOD18 10), SeaDataNet, Coriolis Ocean Dataset, Морского гидрофизического института и Института биологии южных морей, а также других источников. Границы центральной и западной зон высокой солености приняты следующими: 42.5–43.5° с. ш., 30–32.5° в. д. Всего было отобрано 1453 гидрологических станции, выполненных исследовательскими судами в 154 рейсах и семью буями Argo в летний период с 1957 по 2021 г. (июнь – август). Наибольшее ежегодное количество наблюдений относится к 1980-м гг. (до 120 в летний сезон). В последнее время количество летних наблюдений в западной части открытого Черного моря не превышает 20, как в 1960-е гг. Данные каждой экспедиции были проверены на достоверность, заведомо ложные значения (с выбросами значений, не соответствующих аналогичным особенностям в распределении остальных параметров среды) были отбракованы.
На основании этого были рассчитаны средние профили условной плот- ности. Осреднение проводили методом обратных расстояний с последую- щим дополнительным сглаживанием методом низкочастотной фильтрации. Вертикальные профили плотности (формулы EOS-80) были интерполированы с разрешением по глубине 1 м методом, описанным в работе [19], затем был проведен поиск заданного изопикнического уровня на основе минимального отклонения. Перед вычислением изопикнических уровней выполняется кон- троль качества, включая скачки фильтрации и инверсии вертикальной плот- ности. Дальнейшие временны́ е ряды рассчитанных изопикнических уровней
были отфильтрованы с использованием статистического критерия 3σ и затем
усреднены в течение летних периодов каждого года. Процедура усреднения служит определенным фильтром нижних частот для уменьшения внутри- сезонной изменчивости, которая присуща этому слою из-за влияния мезо- масштабных вихрей. Вертикальная дискретизация необработанных данных значительно возросла с 1990-х гг., поэтому эта часть временны́ х рядов имеет более надежные значения изопикнических уровней и проявляет меньше мезомасштабного «шума». Мезомасштабная изменчивость, оцениваемая как стандартные отклонения, рассчитанные в течение летних периодов за последние 30 лет, почти в два раза меньше общей межгодовой изменчивости, что улуч- шает оценки текущего линейного тренда.
Прогнозный анализ проводился с помощью функций прогнозирования MS Excel с использованием AAA-версии алгоритма экспоненциального сгла- живания (ETS), которое в Excel основано на версии AAA (аддитивная ошибка, аддитивный тренд и аддитивная сезонность) алгоритма экспоненциального тройного сглаживания (ETS), который сглаживает незначительные отклоне- ния в тенденциях прошлых данных путем выявления закономерностей сезон- ности и доверительных интервалов. Этот метод прогнозирования лучше всего подходит для нелинейных моделей данных с сезонными или другими повто- ряющимися закономерностями.
Результаты и обсуждение
Изменения структуры водных масс
Усредненные данные о многолетней изменчивости вертикальной струк- туры ХПС во всей глубоководной зоне моря с июня по октябрь в 1993– 2023 гг. приведены на рис. 1. Он показывает, что в 2019 г. верхняя и нижняя границы ХПС объединились на средней глубине около 70 м и эта водная масса, обеспечивающая условия существования реликтовых бореальных видов кор- мового зоопланктона и рыб-планктонофагов ЧМ, исчезла. Этот процесс про- изошел не одномоментно, а в результате постепенного прогрева ядра ХПС за последние 20 лет. Обновление вод его ядра уменьшалось из-за наступ- ления череды теплых зим (рис. 2), которые повторялись примерно через каждые пять лет: в 1996, 2000, 2005, 2010, 2015, 2020 гг. (если оценивать зимы по ТПМ, вычисляемой как среднее за декабрь – февраль).
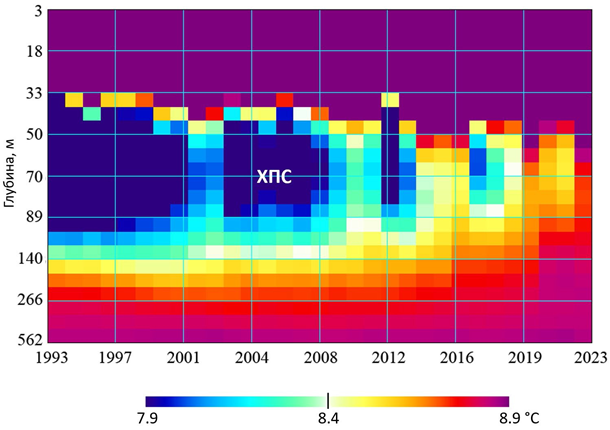
Р и с . 1 . Многолетняя изменчивость ХПС
F i g . 1 . Long-term variability of the cold intermediate layer (CIL)
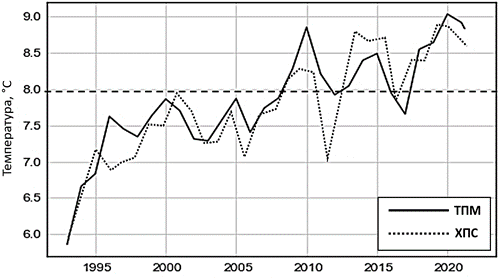
Р ис . 2. Среднезимние значения ТПМ, рассчитан- ные как средние значения за январь, февраль и де- кабрь, и базовое значение ХПС в августе за 1993– 2022 гг.
F ig . 2. Average seasonal (winter) values of sea sur- face temperature, calculated as the average values for January, February and December, and the base value of the CIL in August for 1993–2022
Об этом убедительно говорит высокий коэффициент ранговой корреляции по Спирману [20] между ежегодными значениями ТПМ и ХПС, который ра- вен ~ 0.85 (по Пирсону ~ 0.89). Непараметрический метод ранговой корреля- ции применен здесь, так как ТПМ и ХПС распределены не по нормальному закону согласно тесту Шапиро – Уилка [20]. Смыкание верхней и нижней границ ХПС привело также к повышению солености поверхностного слоя. В 2023 г. на глубине 50 м, которая обычно соответствует положению ядра ХПС, соленость достигла максимального значения 19 ЕПС, что прежде наблюда- лось только в слое 60–70 м, т. е. у среднего положения нижней границы в лет- нее время.
Таким образом, к 2021 г. по всей глубоководной зоне моря установилась новая стратификация водных масс и связанных с ними биотопов. Анализ дан- ных 84 гидрологических профилей по всей глубоководной зоне в слое 0–150 м показал, что под термоклином на всех глубинах вплоть до сероводородной зоны наблюдалась изотермия с температурой около 8.7 °С (рис. 3, а).
Распределение плотностных характеристик воды в этом же слое свиде- тельствовало о том, что СКС и связанный с ним биотоп анаэроконтура нахо- дился в слое 85–115 м. Его верхняя граница соответствовала середине основ- ного пикноклина (σt = 15.8). Средние плотностные границы СКС приводятся в соответствии c [2, 3]. Над ним в кислородной зоне располагалась область обитания планктона и рыб (рис. 3, b) в слое 0–85 м (ВЧВМ и ХПС). Предель- ная глубина нахождения зоопланктона с концентрацией5) 10 мкM, или
-
- мл/л, четко соответствовала верхней границе анаэроконтура по изобате 85 м (рис. 3, c). На глубине 115 м (при плотности воды σt = 16.2) полностью исче- зали следовые концентрации кислорода и начинался слой хемоклина при концентрации H2S около 3 мкМ.

Р ис . 3 . Средний вертикальный профиль температуры (a), условная плот- ность (b) и содержание кислорода (c) в слое 0–150 м в июле – августе 2021 г. в центральных глубоководных районах Черного моря по данным 117-го рей- са ЦКП «НИС Профессор Водяницкий». Синие линии – СКО, черные ли- нии – среднее
F i g. 3. Average vertical profile of temperature (a), density anomaly (b) and oxygen content (c) in the 0–150 m layer in July and August 2021 in the central deep regions of the Black Sea, according to data from the 117th cruise of R/V Pro- fessor Vodyanitsky. Blue lines are SD, black lines are mean values
Изменения структуры биотопов
Из рис. 3, b следует, что зона дневных скоплений батипланктона в 2021 г. соответствовала верхней границе биотопа [12] анаэроконтура, которая нахо- дилась на глубине 85 м. Основанием для исследования многолетней динами- ки вертикального положения анаэроконтура в 1957–2021 гг. послужило то, что среднее положение этой контактной зоны соответствует верхней границе СКС при условной плотности σt = 15.8 [2, 3].
Результаты исследования (рис. 4) свидетельствуют о том, что верхняя граница этого биотопа (σt = 15.8) за 60 лет приблизилась к поверхности на 35 м, а нижняя граница (σt = 16.2) поднялась с глубины 165 до 115 м. Это означает, что в 1983 г. верхняя граница анаэроконтура (белые кружки на рис. 4) поднялась до изобаты 100 м, которая является предельной глубиной обитания черноморского шпрота S. sprattus. С этого момента глубинные скопления холодноводного (кормового) зоопланктона, концентрирующиеся у верхней границы анаэроконтура, оказались в зоне скоплений холодновод- ных рыб-планктонофагов. Подъем нижней границы анаэроконтура (черные кружки на рис. 4) свидетельствует о подъеме хемоклина, т. е. сероводородной зоны, на 50 м вверх.
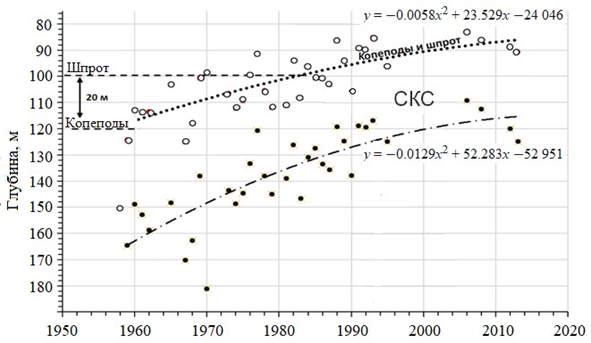
Р ис . 4 . Динамика подъема анаэроконтура к поверхности за 1957–2021 гг. в западно-центральной зоне ЧМ летом: белые кружки – верхняя граница анаэроконтура (σt = 15.8), черные– нижняя граница (σt = 16.2). Усред- ненные данные по годам: пунктирная линия – верхняя контактная зона, штрих-пунктирная ли- ния – нижняя контактная зона; штриховые линии пред- ставляют собой полиномиальные тренды второй степени
F ig . 4. Dynamics of the anaerocontour rise to the surface for 1957–2021, in summer in the western central deep-sea zone of the Black Sea: white circles are the upper anaerocontour boundary (σt = 15.8), black circles are the lower one (σt = 16.2). Average data by year: dotted line – upper contact zone; dashed-dotted line – lower contact zone; dashed lines – sec- ond-degree polynomial trends
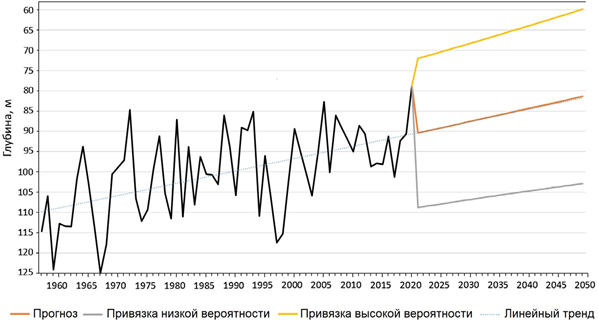
Р ис . 5 . Динамика положения верхней границы СКС до 2050 г. (σt = 15.8) в центре западной части ЧМ (42.5–43.5° с. ш., 30–32.5° в. д.)
F i g. 5. Dynamics of the upper limit of the Suboxic Layer until 2050 (σt = 15.8) in the center of the western Black Sea (42.5–43.5° N, 30–32.5° E)
Зная динамику изменчивости плотностных границ СКС, можно рассчи- тать приблизительное положение границ анаэроконтура на ближайшие 25 лет. Анализ гидрологических данных из центральной части западного циклониче- ского круговорота за 1957–2021 гг. показал: верхняя граница анаэроконтура, регистрируемая по изопикне σt = 15.8 (О2 = 10 мкМ), характеризовалась устой- чивой тенденцией к подъему к поверхности (рис. 5). В 1959 г. среднее по- ложение верхней границы анаэроконтура соответствовало глубине 124 м. В 1982 г. эта контактная зона поднялась на глубину 91 м, то есть почти на 33 м выше (по сравнению с 1959 г.), а в настоящее время верхняя граница СКС в центре западного круговорота расположена на глубине 79 м, что почти на 50 м выше наблюдаемого уровня в 1959 г. Расчет предполагаемых измене- ний положения верхней границы СКС до 2050 г. (пунктирная линия на рис. 5) показал, что через 25 лет она может подняться еще на 9 м.
Обсуждение
Тенденции изменений экологических свойств водных масс
-
-
- ВЧВМ. Установлено, что в конце прошлого века зимняя ТПМ в глубо- ководной зоне моря была около 5.8 °С, сейчас ее среднее значение подня- лось почти до 9 °С (см. рис. 2). Скорость роста ТПМ в ЧМ равна около
-
0.6 °C/10 лет. Это соответствует данным авторов работы [21] о том, что по- вышение ТПМ в 1982 и в 2020 гг. составило 0.40 ± 0.21 и 0.71 ± 0.19 °C/10 лет соответственно. Моделирование [22] показало, что с 1980–1999 по 2080–2099 г. ТПМ повысится еще на 3.7 °C. Согласно нашим расчетам, в последние годы в ВЧВМ стала повышаться соленость: от 18.0–18.4 ЕПС 3) [8] до 18.61–18.8 ЕПС.
-
-
- ХПС. Мы подтверждаем информацию Э. В. Станева с соавторами [15] о том, что к 2019 г. потепление вызвало исчезновение ХПС. В слое воды между серединой сезонного термоклина и серединой пикноклина существен- но изменились основные параметры среды (рис. 1, 6). В период 1993–2023 гг. температура изменилась от 6.0–7.8 до 9 °С, соленость – от 18.0–19.0 до 18.8–
-
20.4 ЕПС, условная плотность σt – от 14.0–14.8 до 14.3–15.8.
-
-
- СКС. За последние 60 лет наблюдается устойчивая тенденция к подъему субкислородного слоя к поверхности (рис. 4, 6). Его верхняя граница (σt = 15.8) в глубоководной зоне моря в 1957 г. была на глубине 115 м, а к 2021 г. она поднялась до 85 м; нижняя граница (σt = 16.2) в 1957 г. была на глубине 165 м, а к 2021 г. она поднялась до 115 м.
- ПЧВМ. Верхняя граница этой водной массы – это хемоклин, от которого вглубь до самого дна простирается сероводородный слой. Результаты настоя- щей работы (рис. 6) свидетельствуют о том, что за 60 лет сероводородный слой поднялся приблизительно на 50 м вверх.
-
Тенденции основных изменений структуры биотопов
- Биотоп поверхностной пленки (нейстон [9]). В настоящее время чис- ленность нейстонных копепод семейства Pontellidae сократилась на 50–70 %. Их встречаемость снизилась настолько, что они были включены в Black Sea Red Book [10]. Дальнейшее повышение ТПМ почти на 4 °C [22] к концу этого сто- летия может не только привести к смене таксономического состава традицион- ных видов нейстона, но и повлиять на биологию организмов мерогипонейстона.
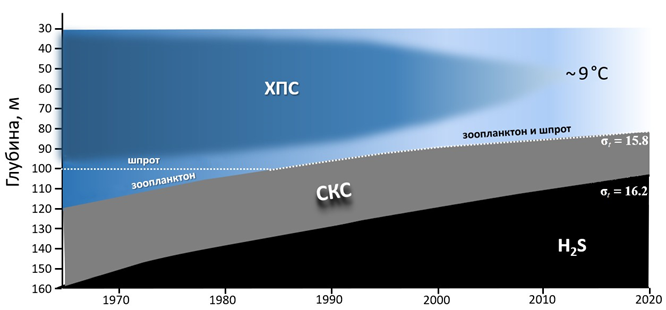
Р ис . 6 . Изменения структуры водных масс, биотопов и верти- кального распределения скоплений зоопланктона и рыб у нижней границы кислородного слоя в центральных глубоководных районах ЧМ в 1960–2020 гг. (днем в летний сезон) в слое 30–160 м
F i g. 6. Changes in the structure of water masses, biotopes and verti- cal distribution of zooplankton and fish aggregations at the lower boundary of the oxygen layer in the central deep waters of the Black Sea in 1960–2020 (day-time, in summer) in the 30–160 m layer
- Биотоп ВЧВМ (эпибионты). Основная масса работ об антропогенных и климатических изменениях экосистемы ЧМ была опубликована преиму- щественно на основе данных об изменениях биоты этого слоя [6, 11, 23, 24]. Поэтому лишь отметим, что в условиях потепления за последние несколько десятилетий биота ВЧВМ начала существенно меняться из-за появления теп- ловодных видов, их преднамеренной или непреднамеренной интродукции. Сейчас в ЧМ зарегистрировано более 300 видов вселенцев, появление кото- рых ускорилось, особенно в последние годы [25]. Сначала было описано 26 но- вых видов вселенцев [26], затем их число возросло до 59. К 2009 г. было опи- сано 156 некоренных видов, причем большинство из них было из Средизем- номорья [27]. В 2017 г. был описан 261 вид вселенцев [28].
- Биотоп ХПС (батибионты). Повышение температуры более 8 °С уже приводит к серьезным нарушениям фенологии нереста умеренно холодно- водных видов рыб, которые начинают нереститься не в декабре, а в конце сентября и заканчивают нерест в мае. В последние годы цикл онтогенетиче- ского развития шпрота S. sprattus от икры до выклева сократился почти в два раза (с 7 до 2–3 дней), уменьшился средний размер личинок при выклеве и их переходе на внешнее питание [29].
Очевидно, что это происходит и с другими холодноводными видами. Известно, что время развития калянуса Calanus euxinus Hulsemann, 1991 при температуре 8 °C составляет до 66 дней (от яйца до взрослой особи), а при температуре 18 °C этот срок сокращается почти в два раза [30]. Сжатие слоя обитания этого вида из-за деоксигенации глубин привело к нарушению трофических связей. В 1960–1970-х гг. стация (объем среды, занятый попу- ляцией [30]) C. euxinus в летнее время занимала всю толщину водной массы
ХПС до глубины 130 м, где находилась изооксигена 10 мкМ [5], в то время как стация черноморского шпрота S. sprattus занимала только верхнюю часть ХПС до глубины 100 м. Черноморский шпрот питается копеподами только днем в зоне их вертикальных миграций или в гипоксийной зоне, где они скапливаются у верхней границы субкислородного слоя.
Чем глубже находится слой концентрации копепод у нижней границы анаэроконтура, тем меньше рыб способно в него опуститься для питания [12]. В 1960–1970-х гг. черноморский шпрот мог перехватывать копепод только во время их суточных вертикальных миграций (рис. 6). В последние годы вся популяция C. euxinus (включая особей в диапаузе) по всей глубоководной зоне ЧМ находится в слое обитания черноморского шпрота. Объединение их стаций 11) произошло в 1985 г. (см. рис. 4), что могло вызвать десятикрат- ный рост запасов шпрота при его максимальной жирности [31]. Именно в эти годы C. euxinus стал преобладать в составе пищевых комков этих рыб. В ближайшей перспективе из-за исчезновения ХПС количество холод- новодного зоопланктона в центральных глубоководных районах может суще- ственно сократиться, поскольку такие виды, как Oithona similis Claus, 1866 и Pseudocalanus elongatus (Brady, 1865) предпочитают температуру менее 8 °C [31].
- Биотоп СКС (анаэроконтур). Изменения положения вертикальных гра- ниц этого биотопа являются главным медиатором климатических изменений экосистемы ЧМ. Результаты настоящего исследования показали, что за по- следние десятилетия верхняя граница анаэроконтура с концентрацией кисло- рода 0.2 мл/л и плотностью σt = 15.8 поднялась почти на 40 м (см. рис. 4). К 2050 г. слой обитания планктона и рыб может сократиться в глубоководных районах еще на 10 м (см. рис. 5).
- Биотоп ПЧВМ. В ближайшие 25 лет в глубоководных районах ЧМ может произойти дальнейший подъем сероводородной зоны к поверхности еще на 10 м (см. рис. 5). В перспективе это может привести к выбросам анаэ- робных вод к поверхности, что может нанести серьезный ущерб экосистеме, рыболовству, рекреационным и биоклиматическим ресурсам побережья Черного моря.
Заключение
Таким образом, результаты настоящего исследования статистически до- стоверно показали, что за последние десятилетия произошли существенные изменения в структуре водных масс и в связанных с ними биотопах гидро- бионтов пелагиали Черного моря. Особенно усложнились условия обитания холодноводных видов в ХПС, который под влиянием потепления смешался с окружающими более теплыми водами. Вполне вероятно, что эта водная масса может вновь появиться под действием многолетних гидрологических циклов и холодноводные организмы не исчезнут.
Огромную роль в этом играет изменчивость интенсивности атмосферной циркуляции над Северной Атлантикой, которая влияет на баланс между по- ступлением высокосоленых водных масс из Средиземного моря (около
170 км3 в среднем за год) и речным стоком в море (около 370 км3 в среднем за год). Этот баланс находится в зависимости от разности уровней между моря- ми, от течений, в том числе дрейфовых, вызываемых ветровым воздействием, а значит может зависеть и от атмосферной циркуляции, изменений климата. Соответственно, и СКС, представленный трансформированной средиземно- морской водой, также связан с изменениями климата.
Изменения объемов речного стока также являются одним из важных следствий климатических изменений, определяющих интенсивность обнов- ления вод ХПС. Установлено, что численность молоди, биомасса и уловы черноморского шпрота S. sprattus находятся в прямой зависимости от дина- мики объемов стока Дуная и Днепра. По-видимому, увеличение стока приво- дит к выносу больших объемов растворенного органического вещества (осо- бенно в районе северо-западного шельфа), обеспечивающих развитие фито- и зоопланктона. Это может создавать благоприятные условия для питания молоди пелагических видов рыб либо приводить к гиперэвтрофикации и об- ширным заморам. Показано, что рост температуры воды ЧМ, по всей вероят- ности, оказывает неблагоприятное влияние на размножение и уловы таких холодолюбивых рыб-хищников, как шпрот, мерланг, ставрида, и придонных бентофагов – палтуса и кефали. Описанный в данной работе процесс исчез- новения ХПС, биотопа обитания холодноводных видов пелагиали ЧМ под влиянием потепления может служить предвестником масштабных экологиче- ских изменений в других регионах Мирового океана.
1. Unexpected changes in the oxic/anoxic interface in the Black Sea / J. W. Murray [et al.] // Nature. 1989. Vol. 338. P. 411–413. https://doi.org/10.1038/338411a0
2. Konovalov S., Murray J., Luther G. Basic processes of Black Sea biogeochemistry // Oceanography. 2005. Vol. 16, iss. 2. P. 24–35. EDN XLNELH. https://doi.org/0.5670/oceanog.2005.39
3. Еремеев В. Н., Коновалов С. К. К вопросу о формировании бюджета и законо- мерностях распределения кислорода и сероводорода в водах Черного моря // Морской экологический журнал. 2006. Т. 5, № 3. С. 5–29. EDN SHTVWD.
4. Investigating hypoxia in aquatic environments: diverse approaches to addressing a complex phenomenon / J. Friedrich [et al.] // Biogeosciences. 2014. Vol. 11, iss. 4. P. 1215–1259. https://doi.org/10.5194/bg-11-1215-2014
5. Decline of the Black Sea oxygen inventory / A. Capet [et al.] // Biogeosciences. 2016. Vol. 13, iss. 4. P. 1287–1297. https://doi.org/10.5194/bg-13-1287-2016
6. Mikaelyan A. S., Zatsepin A. G., Chasovnikov V. K. Long-term changes in nutrient supply of phytoplankton growth in the Black Sea // Journal of Marine Systems. 2013. Vol. 117–118. P. 53–64. https://doi.org/10.1016/j.jmarsys.2013.02.012
7. Видничук А. В., Коновалов С. К. Изменение кислородного режима глубоководной части Черного моря за период 1980–2019 годы // Морской гидрофизический журнал. 2021. Т. 37, № 2. С. 195–206. EDN UMVMXM. https://doi.org/10.22449/0233-7584-2021-2-195-206
8. Мамаев О. И., Архипкин В. С., Тужилкин В. С. T,S-анализ вод Черного моря // Океанология. 1994. Т. 34, № 2. С. 178–192.
9. Зайцев Ю. П. О контурной структуре гидросферы // Гидробиологический журнал. 2015. Т. 51, № 1. С. 3–27.
10. . Abundance and distribution of the hyponeustonic copepods Anomalocera patersonii and Pontella mediterranea in the south-western part of the Black Sea / G. E. Harcotă [et al.] // Turkish Journal of Fisheries and Aquatic Sciences. 2022. Vol. 22, iss. 5. TRJFAS18987. https://doi.org/10.4194/TRJFAS18987
11. Юнев О. A. Коновалов С. К., Великова В. Антропогенная эвтрофикация в пелаги- ческой зоне Черного моря: долгосрочные тенденции, механизмы, последствия. Москва : ГЕОС, 2019. 164 с. EDN YBSYCD.
12. Distribution of Sprattus sprattus phalericus (Risso, 1827) and zooplankton near the Black Sea redoxcline / V. Melnikov [et al.] // Journal of Fish Biology. 2021. Vol. 99, iss. 4. P. 1393–1402. https://doi.org/10.1111/jfb.14848
13. Губарева Е. С., Аннинский Б. Е. Состояние популяции Calanus euxinus (Copepoda) в открытой пелагиали и зоне Крымского шельфа Черного моря осенью 2016 г. // Морской биологический журнал. 2022. Т. 7, № 3. С. 17–27. EDN AONIDN. https://doi.org/10.21072/mbj.2022.07.3.02
14. Мельников В. В. Анаэроконтур Черного моря // Известия Самарского научного центра РАН. 2023. Т. 25, № 5. С. 203–218. EDN NJMWPW. https://doi.org/10.37313/1990-5378-2023-25-5-203-218
15. Stanev E. V., Peneva E., Chtirkova B. Climate change and regional ocean water mass disappearance: Case of the Black Sea // Journal of Geophysical Research: Oceans. 2019. Vol. 124, iss. 7. Р. 4803–4819. https://doi.org/10.1029/2019JC015076
16. Овен Л. С., Шевченко Н. Ф., Володин С. В. Питание шпрота в разных районах Черного моря // Современное состояние ихтиофауны Черного моря : сборник научных трудов. Севастополь : ИнБЮМ им. А. О. Ковалевского, 1996. C. 34–38.
17. Dobricic S., Pinardi N. An oceanographic three-dimensional variational data assimilation scheme // Ocean Modelling. 2008. Vol. 22, iss. 3–4. Р. 89–105. https://doi.org/10.1016/j.ocemod.2008.01.004
18. Strongly coupled data assimilation experiments with linearized ocean-atmosphere balance relationships / A. Storto [et al.] // Monthly Weather Review. 2018. Vol. 146, iss. 4. Р. 1233–1257. https://doi.org/10.1175/MWR-D-17-0222.1
19. Reiniger R. F., Ross C. K. A method of interpolation with applications to oceanographic data // Deep-Sea Research. 1968. Vol. 15, iss. 2. Р. 185–193. https://doi.org/10.1016/0011-7471(68)90040-5
20. Shapiro S. S., Wilk M. B. An analysis of variance test for normality (complete samples) // Biometrika. 1965. Vol. 52, iss. 3–4. Р. 591–611. https://doi.org/10.2307/2333709
21. Mohamed B., Ibrahim O., Nagy H. Sea surface temperature variability and marine heatwaves in the Black Sea // Remote Sensing. 2022. Vol. 14, iss. 10. 2383. https://doi.org/10.3390/rs14102383
22. Climatic controls on biophysical interactions in the Black Sea under present day conditions and a potential future (A1B) climate scenario / H. Cannaby [et al.] // Journal of Marine Systems. 2015. Vol. 141. P. 149–166. https://doi.org/10.1016/j.jmarsys.2014.08.005
23. Oguz T., Dippner J. W., Kaymaz Z. Climatic regulation of the Black Sea hydrometeorological and ecological properties at interannual-to-decadal time scales // Journal of Marine Systems. 2006. Vol. 60, iss. 3–4. P. 235–254. https://doi.org/10.1016/j.jmarsys.2005.11.011
24. Öztürk B. Non-indigenous species in the Mediterranean and the Black Sea. Rome : FAO, 2021. 106 p. (Studies and reviews (General Fisheries Commission for the Medi- terranean) ; no. 87). https://doi.org/10.4060/cb5949en
25. Zaitsev Y., Mamaev V. Marine biological diversity in the Black Sea. A study of change and decline. New York : United Nations Publications, 1997. 208 p. (Black Sea Envi- ronmental Series ; no. 3).
26. Zaitsev Y., Öztürk B. Exotic species in the Aegean, Marmara, Black, Azov and Caspian Seas. Istanbul : Turkish Marine Research Foundation, 2001. 261 p.
27. Shiganova T., Öztürk B. Trend on increasing Mediterranean species arrival into the Black Sea // Climate forcing and its impacts on the Black Sea marine biota / ed. by F. Briand. Monaco : CIESM, 2009. P. 75–91. (CIESM Workshop Monographs ; no. 39).
28. Alexandrov B., Minicheva G., Zaitsev Y. Black Sea network of Marine Protected Areas: European approaches and adaptation to expansion and monitoring in Ukraine // Management of Marine Protected Areas: A Network Perspective / ed. P. D. Goriup. Chichester : John Wiley and Sons, 2017. P. 227–246. https://doi.org/10.1002/9781119075806.ch12
29. Влияние некоторых абиотических и биотических факторов на нерест европейского шпрота Sprattus sprattus (Linnaeus, 1758) в Черном море в ноябре 2016–2017 гг. / Т. Н. Климова [и др.] // Океанология. 2021. Т. 61, № 1. С. 67–78. EDN YEQEHK. https://doi.org/10.31857/S0030157421010081
30. Сажина Л. И. Рост массовых копепод Черного моря в лабораторных условиях // Биология моря. 1973. Т. 28. С. 41–51.
31. Воздействие глобальных климатических и региональных факторов на мелких пелагических рыб Черного моря / Г. Е. Шульман [и др.] // Морской экологиче- ский журнал. 2007. Т. 6, № 4. С. 18–30. EDN UIQKNH.