












Russian Federation
Russian Federation
Purpose. The aim of the study is to analyze the measurement data on oxygen concentration at the bottom of Amur Bay in the cold season, and to reveal the reasons for its decrease in the bottom layer in early spring (late February – March) when the bay is still covered with ice. Methods and Results. The variations in bottom water characteristics during the cold season were studied in the area of summer hypoxia at the 22 m depth (1 m from the seafloor) using the autonomous bottom station Water Quality Monitor from Wet Labs. Temperature, salinity (by conductivity), dissolved oxy-gen (DO), and chlorophyll a (fluorescence) were recorded every 4 hours. The monitoring data for the cold period 2013–2014 were compared to the data obtained previously at the same station and at the site with the same coordinates in summer 2011. The basic patterns of changes in oxygen content, and the periods of production dominance and organic matter mineralization in the bottom layer of Amur Bay in the cold season were determined. Conclusions. During the winter season, upwelling of the Japan Sea waters delivers nutrients to the Amur Bay bottom layer. Due to the vertical mixing conditioned by low stability of waters the euphotic layer is supplied with nutrients, and photosynthesis takes place in the entire water column of the bay. During four months in winter, the Amur Bay waters become oversaturated with oxygen as compared to its content in the atmosphere. The onset of summer monsoon (late February – early March) marks the beginning of formation of summer hypoxia in the Amur Bay bottom waters.
Japan Sea, Amur Bay, dissolved oxygen, hypoxia, upwelling, downwelling, nutrients, photosynthesis
Введение
Деоксигенация Мирового океана и его шельфа относится к глобальной проблеме, которая является предметом интенсивного исследования. Существует общее согласие в отношении причин образования гипоксии/аноксии на шельфе Мирового океана, главная из которых – эвтрофикация прибрежных вод [1]. В заливе Петра Великого (ЗПВ) расположены две впадины (рис. 1), в которых в летний сезон формируется гипоксия (содержание кислорода менее 76 мкмоль/кг). Одна впадина находится у о. Фуругельма (юго-западная часть залива), в ней гипоксия носит нерегулярный характер, а вторая, в которой регистрируется сезонная гипоксия, – в центральной части Амурского залива [2]. Основная причина образования гипоксии – поступление биогенных веществ в период паводков эвтрофированными реками Туманной и Раздольной [2].
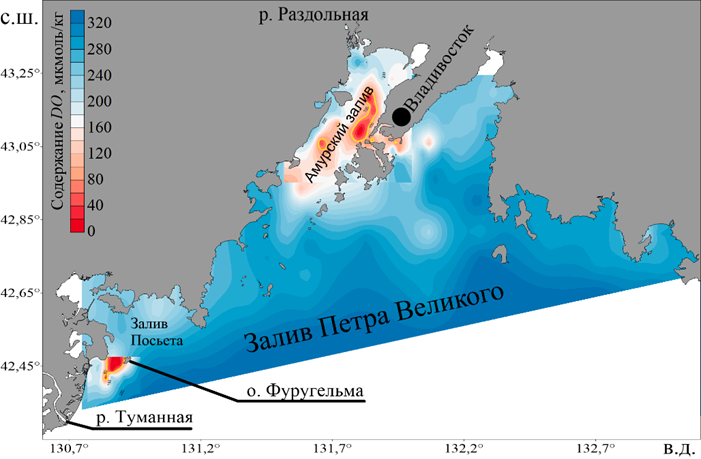
Р и с. 1. Расположение областей гипоксии в заливе Петра Великого
F i g. 1. Hypoxia areas in the Peter the Great Bay
Амурский залив представляет собой часть ЗПВ. Географически он вытянут с северо-востока на юго-запад. Протяженность залива 70 км, ширина колеблется от 10 до 22 км [3]. Максимальные глубины Амурского залива не превышают 53 м на крайнем южном участке. Средняя глубина залива около 30 м [4]. В северной части залива находится обширная мелководная зона с глубинами, не превышающими 10 м. Центральную часть акватории занимает глубоководная впадина, достигающая глубины 20–25 м. На юго-востоке эта впадина ограничивается Муравьевским порогом, протянувшимся от п-ова Песчаного в юго-восточном направлении вплоть до о. Русского (рис. 2). В летнее время во впадине формируется гипоксия придонного слоя вод, верхняя граница которой залегает на глубинах 15–17 м [5]. Под гипоксией понимают состояние экосистемы с таким низким содержанием кислорода в воде, при котором происходят количественные и качественные изменения экосистемы. Литературные источники предлагают различные пороговые значения концентрации кислорода в качестве критерия гипоксии. Чаще всего они находятся в диапазоне от 63 до 89 мкмоль/кг [6]. В нашей работе в качестве критерия гипоксии используется среднее значение 76 мкмоль/кг.
Муссонный климат Приморья [7] обеспечивает развитие даунвеллинговой циркуляции вод Амурского залива в весенне-летний сезон и апвеллинговой – в осенне-зимний сезон. Даунвеллинг способствует формированию гипоксии, а апвеллинг – ее разрушению [8].
В теплый сезон было проведено много гидрохимических исследований Амурского залива, которые были направлены на установление особенностей формирования и разрушения гипоксии. Обзор этих работ можно найти в [6]. Гораздо в меньшей степени изучены гидрохимические процессы в зимний сезон. Результаты наших исследований указывают на высокие концентрации кислорода зимой (400–500 мкмоль/кг) в основной части залива как в поверхностном, так и в придонном слое [5]. Более того, отмечается пересыщение кислородом придонных вод Амурского залива относительно атмосферного уровня, за исключением Тавричанского лимана и залива Углового В зимний сезон наблюдается достаточно интенсивная первичная продукция подо льдом (0,1–0,3 гС/м2) [9]. В то же время обширные гидрологические исследования, которые включали измерения содержания кислорода и хлорофилла зондом, дали основание авторам работы [10] сделать предположение о возможности формирования гипоксии придонных вод Амурском заливе в зимний сезон. В данной работе мы приводим результаты мониторинговых наблюдений, полученные в холодное время года заякоренной станцией, которая была помещена в область летней гипоксии Амурского залива.
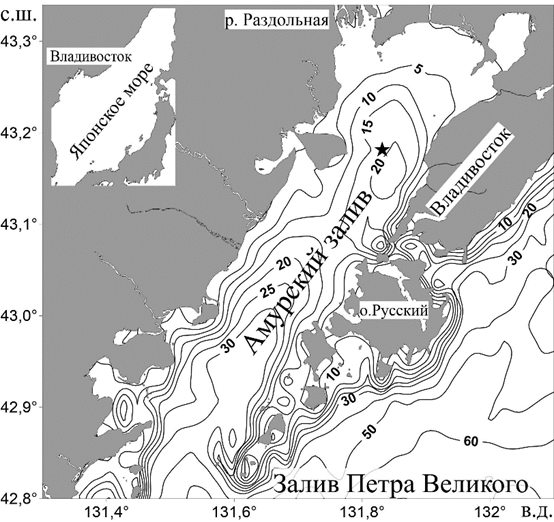
Р и с. 2. Географическое расположение Амурского залива и карта глубин. Звездочкой отмечено расположение мониторинговой станции WQM
F i g. 2. Geographical location of Amur Bay, and depth map. Star indicates the WQM station location
Цель работы – изучить изменения параметров воды и установить основные причины изменения содержания кислорода у дна в холодное время года.
Методы измерений
Изменчивость характеристик придонной воды в холодное время года была изучена в районе, в котором летом регистрируется гипоксия, на глубине 22 м, на расстоянии 1 м от дна с помощью автономной придонной станции Water Quality Monitor (WQM) фирмы Wet Labs. Координаты местоположения станции 43°10,894′ с. ш., 131°49,949′ в. д. (рис. 2). Измерения проводили с 11 сентября 2013 г. по 15 мая 2014 г. Каждые 4 часа велась регистрация температуры, солености (по электропроводности), концентрации растворенного кислорода (Dissolved Oxygen, сокращ. DO) и хлорофилла а (флуоресценция) с интервалом в 1 с в течение 5 мин подводным логгером DH4 той же фирмы. Для мониторинговой станции WQM компанией WET Labs решены проблемы биообрастания благодаря использованию медных корпусов, инжекции хлорной извести и вкладышам, содержащим пестициды. Такие меры обеспечили стабильную работу датчиков в течение длительного периода времени [11]. В цитируемой выше статье даны подробные характеристики датчиков WQM. Разница между результатами измерений кислорода датчиком станции WQM и измерениями по методу Винклера составляла 1,2–14,0 мкмоль/кг [8]. Наиболее высокое расхождение в результатах мы связываем с несоответствием расположения станции WQM и батометра в процессе пробоотбора. Данные мониторинга для холодного периода 2013/14 г. сравнивали с данными для летнего сезона 2011 г., ранее полученными этой же станцией, расположенной в точке с почти идентичными координатами (43°10,881′ с. ш., 131°49,893′ в. д.) [8].
Летнее и зимнее состояние вод Амурского залива мы сравнивали, используя кажущееся поглощение кислорода (Apparent Oxygen Utilization, сокращ. AOU), поскольку этот параметр характеризует баланс между продукционными и деструкционными процессами. Значения AOU рассчитывали с использованием соотношения

Здесь AOU рассчитывали как разницу между концентрацией кислорода в морской воде, находящейся в равновесии с атмосферой ([O2]o), при заданной температуре и солености и измеренной концентрацией ([O2]). Растворимость кислорода в морской воде при заданной температуре и солености рассчитывали по уравнению, представленному в работе [12]. Очевидно, что морская вода находится в состоянии избыточного содержания кислорода по отношению к атмосферному уровню при AOU < 0 и дефицита, когда AOU > 0.
В ранневесенний сезон (22 марта 2011 г.) была выполнена гидрохимическая станция в области летней гипоксии (43°11,816′ с. ш., 131°50,139′ в. д.). Станция была выполнена со спасательного судна в период таяния льда, когда северная часть Амурского залива оставалась покрыта битым льдом, а южная была свободна ото льда. Были получены вертикальные профили парциального давления СО2 (рСО2), концентраций кислорода, аммонийного азота, фосфатов и силикатов в области летней гипоксии. Щелочность измеряли прямым титрованием в открытой ячейке соляной кислотой (0,02N) со смешанным индикатором (метиловый красный + метиленовый голубой) Измерения рН выполняли при температуре 20 °С потенциометрическим методом в ячейке без жидкостного соединения электродов [13]. Расчет парциального давления СО2 проводили в соответствии с [14]. Измерения биогенных веществ выполняли по стандартным методикам
Результаты
Детальная изменчивость гидрологических характеристик придонных вод Амурского залива в области формирования гипоксии для холодного периода 2013/14 г. представлена на рис. 3. В качестве дополнительной информации для демонстрации полного внутригодового цикла привлечены результаты работы станции WQM, полученные в 2011 г. и подробно рассмотренные ранее [8].
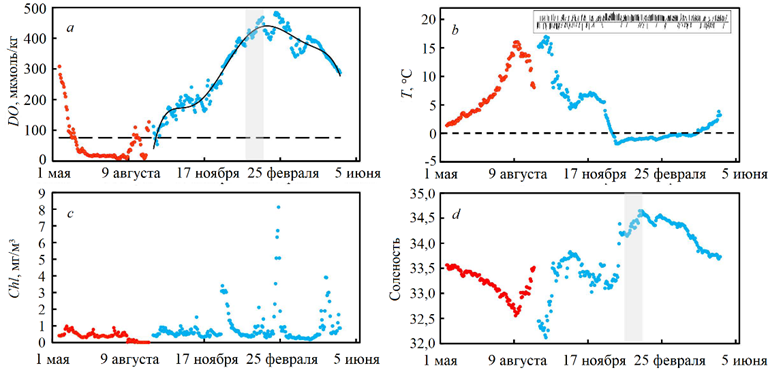
Р и с. 3. Временна́я изменчивость концентрации растворенного кислорода (штриховая линия обозначает уровень гипоксии – 76 мкмоль/кг) (a), температуры (b), содержания хлорофилла а (c) и солености (d) в придонном слое вод Амурского залива в точке расположения автономной придонной станции WQM: в 2011 г. (●); в 2013/14 г. (●). На врезке рис. 3, b представлена временна́я изменчивость направления ветра в 2013/14 г. Серая область соответствует периоду 27 декабря – 02 февраля, когда происходит увеличение солености вследствие образования рассолов
F i g. 3. Temporal variability of dissolved oxygen concentration (dashed line indicates the hypoxia level – 76 μmol/kg) (a), temperature (b), chlorophyll a content (c) and salinity (d) in the Amur Bay bottom layer at the location of autonomous bottom station WQM: in 2011 (●); in 2013–2014 (●). The inset in Fig. 3, b shows temporal variability of wind direction in 2013–2014. Grey area corresponds to the period December 27 – February 02 when salinity increases due to brine formation
В конце августа – осенью в результате апвеллинга япономорских вод происходит разрушение придонной гипоксии [8], что находит свое отражение в росте DO, отсутствии хлорофилла, уменьшении температуры воды и увеличении солености. Изменение концентрации нитрат-ионов, отсутствующих летом в придонном слое воды Амурского залива, свидетельствует о подъеме япономорской воды 1 с глубины 250–300 м. Для этих глубин концентрации силикатов, нитратов, фосфатов и растворенного кислорода равны 29,3; 17,2; 1,36; 285 мкМ соответственно [15]. В 2013 г. вытеснение бескислородных вод из рассматриваемой области холодными солеными япономорскими водами происходило с 12 сентября по 24 октября вследствие осеннего апвеллинга (рис. 3). Процесс сопровождался снижением температуры воды с 17,0 до 4,3 °С. Соленость в этот период возрастала с 32,12 до 33,69, а DO – с 54 до 213 мкмоль/кг. В этот период наблюдений направление ветра было неустойчивым, менялось с южного на северное и обратно (врезка на рис. 3, b).
Мы полагаем, что период с 25 октября по 09 декабря следует рассматривать как продолжение перехода с летнего муссона на зимний. Этот период характерен меняющимся направлением ветра, что приводит к изменениям в циркуляции вод залива и разнонаправленным во времени изменениям наблюдаемых параметров: DO в диапазоне 159–263 мкмоль/кг; температуры – 4,6–7,2 °C; солености – 33,03–33,79. Существенные вариации в этот переходный период вызваны не только сменой направления ветра, но и ветровым перемешиванием япономорской воды с остаточной шельфовой водой летнего происхождения, а также атмосферным охлаждением воды, которое сопровождалось растворением атмосферных газов. Рост концентрации кислорода в этот период отчасти обусловлен увеличением растворимости газов при понижении температуры, а также ростом первичной продукции.
На побережье Амурского залива 2013 г. 10 декабря выпал снег и началось интенсивное ледообразование в заливе. Температура в заливе опустилась до точки замерзания морской воды, −1,818 °С (27 декабря). С небольшим запозданием начали повышаться соленость, до 33,323, и концентрация кислорода, до 353 мкмоль/кг. В этот период (17 декабря) наблюдался максимум хлорофилла – 3,4 мг/м3 (рис. 3, c). В период с 10 по 27 декабря происходило зимнее охлаждение, которое сопровождалось формированием сплошного ледяного покрова в северной части Амурского залива (к северу от широты 43°8,5′ с. ш. до устья р. Раздольной) и зимней конвекцией. Ниже параллели 43°8,5' с. ш. также наблюдалось образование льда, однако оно не достигало сплошного. Доминирование ветров северных направлений усиливало этот эффект. Среднесуточная температура воздуха 8 декабря 2013 г. составила 1,15 °С, 9 декабря – 5,51 °С (http://www.rp5.ru/). По данным, взятым с сайта https://worldview.earthdata.nasa.gov, к 11 декабря северная часть Амурского залива была покрыта сплошным льдом, установка WQM находилась подо льдом. Тот факт, что 27 декабря среднесуточная температура воды у дна достигла точки замерзания, означает однородность параметров воды по вертикали во всей толще вод залива, поскольку замерзание воды (ледообразование) происходит на поверхности залива. С 27 декабря по 02 февраля наступает период, характеризующийся ростом солености (до 34,578) и DO (до 468 мкмоль/кг), на рис. 3, a и d этот период обозначен серым цветом. В это же время наблюдается небольшое увеличение температуры воды (до −0,878 °С). Максимальная соленость япономорских вод равна 34,070 ± 0,002 [15], а соленость вод Амурского залива в теплый сезон существенно меньше этого значения (рис. 3, d) [3]. Поэтому увеличение солености до 34,578 являлось результатом образования рассолов при льдообразовании. Небольшое увеличение температуры воды мы связываем с нагревом воды от морского дна, которое частично хранит тепло летнего сезона [16, 10]. Подогрев воды снизу приводит к интенсификации вертикального перемешивания. Однородность толщи воды сохраняется, но с незначительно меняющимися T, S-по-казателями во времени (рис. 3, b, d).
Февраль характеризовался незначительными колебаниями гидрологических характеристик. С 2 по 12 февраля наблюдалось незначительное уменьшение DO (до 396 мкмоль/кг), увеличение температуры (до −0,69 °С) и понижение солености (до 34,413). С 12 февраля по 23 февраля, напротив – отмечалось увеличение DO (до 477 мкмоль/кг), понижение температуры воды (до −0,856 °С) и рост солености (до 34,56). Наиболее вероятной причиной этих колебаний может быть адвекция вод в заливе, а увеличение концентрации DO обусловлено «цветением» фитопланктона. Максимум концентрации хлорофилла – 8 мг/м3 (23 февраля) – соответствовал максимуму концентрации DO (рис. 3).
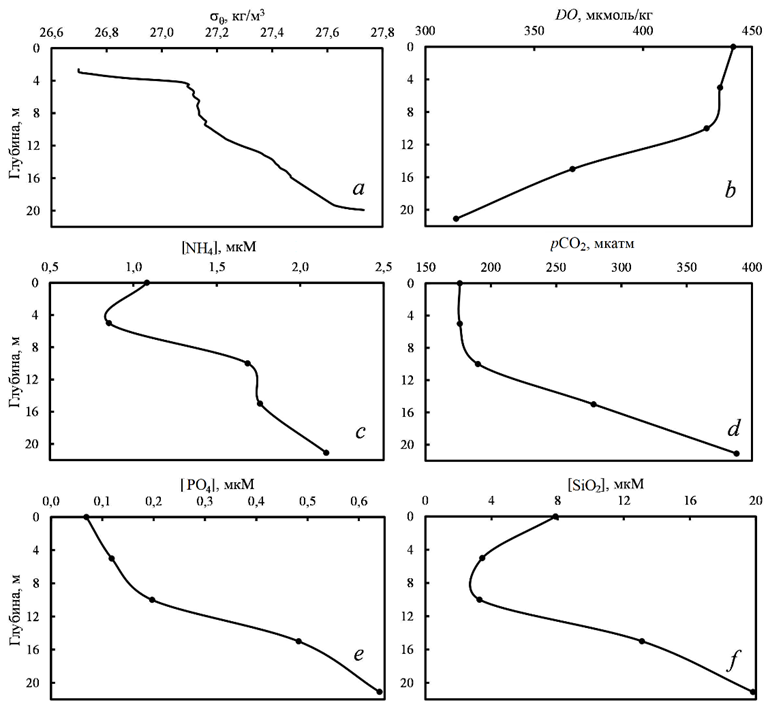
Р и с. 4. Вертикальные профили условной плотности (a), концентрации кислорода (b), концентрации аммонийного азота (c), парциального давления углекислого газа (d), концентраций фосфатов (e) и силикатов (f) в области формирования гипоксии (43°11,816′ с. ш., 131°50,139′ в. д.), Амурский залив, 22 марта 2011 г.
F i g. 4. Vertical profiles of conditional density (a), oxygen concentration (b), ammonium nitrogen concentration (c), partial pressure of carbon dioxide (d), and phosphate (e) and silicate (f) concentrations in the area of hypoxia formation (43°11.816′ N, 131°50.139′ E) in Amur Bay, March 22, 2011
Формирование вертикальной гидрологической и гидрохимической стратификации вод Амурского залива наблюдается в марте (рис. 4), оно обусловлено таянием льда и изменением циркуляции вод с апвеллингового типа на даунвеллинговый, вызванным изменениями направления ветра (см. рис. 3, b) [8].
Обсуждение
Муссонный климат Приморья играет важную роль в формировании и разрушении гипоксии придонных вод Амурского залива. Юго-восточное направление ветра в летний сезон формирует даунвеллинговую циркуляцию в заливе [8]. Даунвеллинговая циркуляция ограничивает взаимодействие подповерхностных вод Японского моря с водами шельфа ЗПВ. В этот сезон выпадает основное количество атмосферных осадков [7]. Поток эвтрофированных вод р. Раздольной в Амурский залив летом существенно возрастает. Сочетание этих двух факторов создает условия, при которых основным источником биогенных веществ в залив является речной сток, что вызывает «цветение» фитопланктона в поверхностных водах залива. Следует отметить, что выделившийся при фотосинтезе кислород частично уходит в атмосферу, а частично остается в верхнем слое залива вследствие устойчивой стратификации. Часть образовавшейся избыточной биомассы фитопланктона (не съеденной зоопланктоном и зообентосом) поступает на дно залива. Во впадинах залива, ниже глубин 15–17 м, куда не проникает ФАР (фотосинтетически активная радиация), формируется гипоксия в результате микробиологического разложения биомассы фитопланктона [2].
В холодный период года северо-западное направление ветра формирует апвеллинговую циркуляцию вод залива. В это время резко сокращается выпадение атмосферных осадков [7]. Расход р. Раздольной в зимний сезон уменьшается более чем в 100 раз в сравнении с летними паводками. В то же время подповерхностная вода Японского моря, поднимающаяся с глубины 250–300 м, содержит достаточно высокие концентрации биогенных веществ и растворенного кислорода: силикатов – 29,3, нитратов – 17,2, фосфатов – 1,36 мкМ и растворенного кислорода – 285 мкмоль/кг [15]. Таким образом, в зимний сезон основным источником биогенных веществ, поставляемых в Амурский залив, служат подповерхностные воды Японского моря. Эти воды прозрачные, поэтому ФАР проникает до дна в свободной от сплошного льда южной части залива. Фотосинтез активно протекает и подо льдом, если он не покрыт снегом. Кроме того, эти воды поступают в придонный слой залива, а потом, благодаря зимней конвекции, перемешиваются. Биогенные вещества, поступающие из Японского моря, обеспечивают зимний фотосинтез вод Амурского залива и, соответственно, высокое содержание кислорода (см. рис. 2). Используя стехиометрию Рэдфилда [17], фотосинтез органического вещества можно записать схемой
Мы полагаем, что япономорские воды в результате апвеллинга полностью заменяют шельфовые воды Амурского залива. Если в этих водах происходит фотосинтез, то концентрация DO будет составлять (285 + 138 × [PO4]) = 473 мкмоль/кг, что сравнимо с максимальной концентрацией DO в зимний сезон – 482 мкмоль/кг (см. рис. 3, а). Дополнительным источником поступления биогенных веществ в залив является поток биогенных веществ от дна [18].
Сравнение AOU для теплого и холодного сезонов показано на рис. 5. Из него видно, что летом в условиях даунвеллинговой циркуляции вод Амурского залива в придонных водах доминируют деструкционные процессы. Значения AOU положительные и достигают 300 мкмоль/кг (рис. 5). Проникновение япономорских вод в залив осенью существенно снижает это доминирование, поскольку привносит кислород в придонные воды залива. Однако смена знака продукционно-деструкционного баланса, несмотря на благоприятные кислородные условия, происходит лишь 29 декабря. Это объясняется тем, что япономорские воды приходят с глубины 250–300 м, где отсутствует фотосинтез, и эти воды также испытывают дефицит кислорода по отношению к атмосферному уровню. Сравнение рис. 3, a и 5 приводит нас к выводу, что смена продукционно-деструкционного баланса наступает лишь вследствие интенсификации процессов формирования органического вещества в декабре, которые усиливаются в результате поставки биогенных веществ япономорскими водами и по времени совпадают с условиями зимней конвекции, когда весь вертикальный столб воды залива достигает температуры замерзания. Из рис. 5 следует, что придонные воды залива пересыщены по отношению к атмосферному кислороду в течение почти четырех месяцев (29.12.2013–22.04.2014).
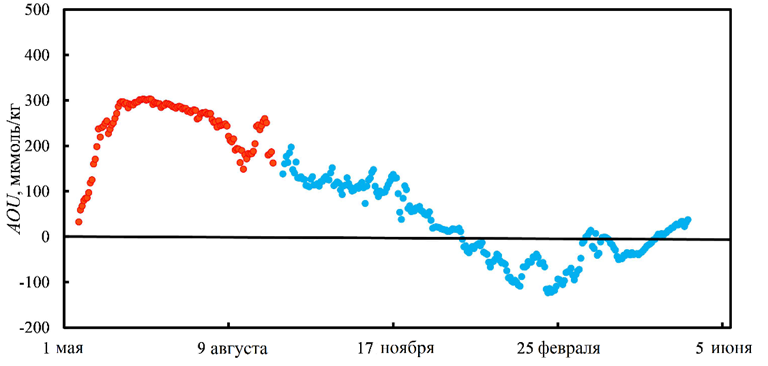
Р и с. 5. Временна́я изменчивость AOU у дна Амурского залива в точке расположения станции WQM: в 2011 г. (●); в 2013/14 г. (●)
F i g. 5. Temporal variability of AOU in the Amur Bay bottom layer at the location of WQM station: in 2011 (●); in 2013–2014 (●)
Придонные воды в точке расположения станции WQM весь зимний сезон характеризуются высоким содержанием DO, в 4–6 раз превышающим пороговое значение 76 мкмоль/кг, ниже которого состояние вод характеризуется как гипоксийное. Одна из главных причин состоит в том, что, в отличие от летнего сезона, поставка биогенных веществ в придонные воды залива зимой происходит в условиях высокой прозрачности япономорских вод и отсутствия их плотностной стратификации. Поэтому в зимних условиях фотосинтез, благодаря вертикальному перемешиванию, затрагивает всю не покрытую льдом водную толщу и в придонный слой поступает вода с AOU < 0, т. е. пересыщенная по отношению к атмосферному уровню. Зимнее «цветение» фитопланктона возможно и подо льдом. Объяснение было предложено в работе [19]. Покров льда уменьшает интенсивность ФАР. Однако фотосинтез подо льдом в мелководных бассейнах, таких как бух. Новгородская (глубина 10–15 м), зал. Посьета, возможен благодаря зимней конвекции. В условиях зимней конвекции происходит вертикальное перемешивание всей толщи воды в мелководной бухте. Биогенные вещества поставляются в подледный слой, где происходит фотосинтез. Клетки фитопланктона увлекаются конвекцией в более глубокие слои, а потом возвращаются вновь в область фотосинтеза. Таким образом, малая глубина бассейна препятствует выносу клеток фитопланктона из зоны влияния ФАР. На наш взгляд, ситуация, изложенная в приведенной ранее публикации, относится именно к участкам Амурского залива, занятым льдом при отсутствии снежного покрова.
Наблюдаемое снижение уровня кислорода после 23 февраля свидетельствует о наличии общих механизмов, ведущих к уменьшению содержания кислорода в водах залива. На наш взгляд, таких процессов два. В зимний сезон, когда AOU < 0, газообмен в акватории залива, не покрытой льдом, будет приводить к уменьшению концентрации кислорода. Такая площадь возрастает в конце февраля – начале марта, и к началу апреля залив полностью очищается от льда (URL: https://worldview.earthdata.nasa.gov). Такой механизм извлечения кислорода из вод залива функционирует эффективно лишь тогда, когда концентрация растворенного кислорода превышает атмосферный уровень, т. е. до 26 апреля (рис. 5). Однако и после перехода экосистемы залива от преобладания продукционных процессов (AOU < 0) к превалированию деструкционных процессов (AOU > 0), содержание DO в придонном слое залива продолжает снижаться (см. рис. 3, 5). Второй процесс, приводящий к уменьшению DO, состоит в биохимическом потреблении кислорода при минерализации органического вещества на границе раздела вода – дно. Мы использовали ранее полученную скорость биохимического потребления кислорода VБПК (мкмоль/(л×сут)) придонным слоем Амурского залива [8]:
VБПК = −0,1225×[O2].
Результаты наблюдений за концентрацией кислорода в зависимости от времени (см. рис. 2) были обработаны методом наименьших квадратов. Было получено эмпирическое уравнение для [O2] (мкмоль/л):
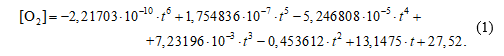 (1)
(1)
Здесь t – время в сутках от начала наблюдений. Производная от зависимости (4) представляет собой общую скорость изменения содержания кислорода Vобщ в любой момент времени. Скорость биохимического окисления кислорода VБПК у дна рассчитывали из соотношения (1), а скорость вентиляции Vвент вод – как разность между Vобщ и VБПК. Результаты расчетов представлены на рис. 6. Биохимическое потребление кислорода является реакцией второго порядка [8]. Иначе говоря, скорость потребления кислорода максимальная при максимальной концентрации DO (рис. 6; уравнение (1)).
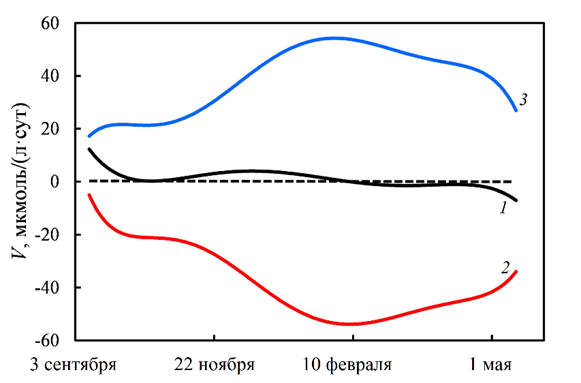
Р и с. 6. Зависимость скорости изменения концентрации кислорода от времени: 1 – общая скорость Vобщ, рассчитанная по уравнению (1); 2 – скорость биохимического потребления на окисление органического вещества VБПК; 3 – скорость вентиляции вод Vвент
F i g. 6. Dependence of rate of oxygen concentration change on time: 1 – total rate Vtot calculated by equation (1); 2 – rate of biochemical consumption for oxidation of organic matter VBCO; 3 – rate of water ventilation Vvent
В марте 2011 г. были получены профили гидрохимических параметров (см. рис. 4). На момент исследования (22 марта) возникла плотностная стратификация вод из-за таяния льда и сезонного повышения температуры. В этот период происходит смена апвеллинговой циркуляции вод залива на даунвеллинговую. Даунвеллинговая циркуляция ограничивает воздействие Японского моря на Амурский залив. Поэтому рассматриваемые профили на рис. 4 обусловлены геохимическими процессами у дна залива. Концентрация DO ко дну уменьшалась, а концентрации биогенных веществ возрастали, что указывает на минерализацию органического вещества на границе раздела вода – осадок. Содержание органического углерода в донных осадках в местах гипоксии составляет 2–3 % [20]. Его минерализация внутри осадка происходит по анаэробному пути. Даунвеллинг приводит к ресуспензии поверхностного осадка на дне залива, что увеличивает мутность придонных вод и поток продуктов минерализации органического вещества из донных осадков в воду [8]. Поэтому, несмотря на достаточно высокие концентрации кислорода, основным продуктом минерального азота был аммоний (см. рис. 4, c). Концентрации нитрат-ионов также измерялись, однако их содержание было примерно в 2–3 раза меньше в сравнении с аммонием. Мы не исключаем и другой путь, когда на границе раздела вода – осадок в случае восстановленных илов формируется слой с низким содержанием кислорода [21]. Тогда в тонком слое с низким содержанием кислорода минерализация органического вещества также будет приводить к образованию аммония, а не нитрат-ионов [6]. Этот вопрос требует дополнительных исследований.
Следует отметить важность переходного периода: в конце февраля – начале марта происходит смена направления ветра с зимнего муссона на летний, что приводит к изменению апвеллинговой циркуляции вод залива на даунвеллинговую; в этот период ослабевает взаимодействие Японского моря с шельфом и, соответственно, поставка биогенных веществ япономорскими водами. Интенсивность фотосинтеза в заливе снижается; совокупность этих факторов, по сути, служит началом формирования летней гипоксии.
Выводы
Изменение содержания кислорода у дна Амурского залива определяется совокупным влиянием динамики вод и биогеохимических процессов.
Существуют два основных природных источника биогенных веществ, которые определяют направленность биогеохимических процессов в Амурском заливе и его состояние: 1) эвтрофированные воды р. Раздольной; 2) япономорские воды с глубины 250–300 м.
В холодное время года апвеллинг япономорских вод поставляет биогенные вещества в придонный слой залива. Вертикальное перемешивание, обусловленное низкой статической устойчивостью вод, обеспечивает биогенными веществами эвфотический слой. Фотосинтез, благодаря вертикальному перемешиванию, затрагивает всю толщу вод залива. Зимние воды Амурского залива пересыщены относительно атмосферного уровня в течение четырех месяцев, при этом концентрация достигает 353–482 мкмоль/кг.
Смена функционирования экосистемы Амурского залива происходит дважды в год в связи с изменением типа циркуляции вод в заливе: в феврале – марте возникает даунвеллинговый тип циркуляции, который инициирует поставку биогенных веществ из р. Раздольной, формирование вертикальной стратификации вод и уменьшение содержания кислорода у дна, летом достигающего критических значений менее 76 мкмоль/кг; в сентябре – октябре формируется апвеллинговый тип циркуляции, который разрушает летнюю гипоксию и поставляет биогенные вещества из подповерхностных вод Японского моря.
1. Declining oxygen in the global ocean and coastal waters / D. Breitburg [et al.] // Science. 2018. Vol. 359, no. 6371. eaam7240. http://dx.doi.org/10.1126/science.aam7240
2. Impact of the transboundary Razdolnaya and Tumannaya Rivers on deoxygenation of the Peter the Great Bay (Sea of Japan) / P. Ya. Tishchenko [et al.] // Estuarine, Coastal and Shelf Science. 2020. Vol. 239. 106731. http://dx.doi.org/10.1016/j.ecss.2020.106731
3. Gomoyunov K. A. Gidrologicheskiy ocherk Amurskogo zaliva i reki Suyfuna. Vladivo-stok : tip. Gos. Dal'nevost. un-ta, 1926. 44 s. (Trudy Gosudarstvennogo Dal'nevostochno-go universiteta. Seriya 3 ; 1).
4. Grigor'ev R. V., Zuenko Yu. I. Srednemnogoletnee raspredelenie temperatury i soleno-sti v Amurskom zalive Yaponskogo morya // Izvestiya TINRO. 2005. T. 143. S. 179–188. EDN HSLAUN.
5. Sezonnaya gipoksiya Amurskogo zaliva (Yaponskoe more) / P. Ya. Tischenko [i dr.] // Izve-stiya TINRO. 2011. T. 165. S. 136–157. EDN OEUVNH.
6. Gipoksiya zaliva Petra Velikogo / P. P. Tischenko [i dr.] // Izvestiya TINRO. 2021. T. 201, № 3. S. 600–639. EDN SNBBPO. https://doi.org/10.26428/1606-9919-2021-201-600-639
7. Gayko L. A. Osobennosti gidrometeorologicheskogo rezhima pribrezhnoy zony zaliva Petra Velikogo (Yaponskoe more). Vladivostok : Dal'nauka, 2005. 150 s.
8. Summertime in situ monitoring of oxygen depletion in Amursky Bay (Japan/East Sea) / P. Tishchenko [et al.] // Continental Shelf Research. 2016. Vol. 118. P. 77–87. https://doi.org/10.1016/j.csr.2016.02.014
9. Sorokin Yu. I., Konovalova I. W. Production and decomposition of organic matter in a bay of the Japan Sea during the winter diatom bloom // Limnology and Oceanography. 1973. Vol. 18, iss. 6. P. 962–967. https://doi.org/10.4319/lo.1973.18.6.0962
10. Lazaryuk A. Yu., Lobanov V. B., Ponomarev V. I. Evolyuciya termohalinnoy struktury vod Amurskogo zaliva v holodnyy sezon // Vestnik Dal'nevostochnogo otdeleniya Rossiy-skoy akademii nauk. 2013. № 6 (172). S. 59–70. EDN THYCTX.
11. WQM: a new integrated water quality monitoring package for long-term in-situ observation of physical and biogeochemical parameters / S. M. Orrico [et al.] // Proceedings of OCEANS 2007, Vancouver, BC, Canada, 2007. P. 1–9. https://doi.org/10.1109/OCEANS.2007.4449418
12. Weiss R. F. The solubility of nitrogen, oxygen and argon in water and seawater // Deep Sea Research and Oceanographic Abstracts. 1970. Vol. 17, iss. 4. P. 721–735. https://doi.org/10.1016/0011-7471(70)90037-9
13. Izmerenie rN v estuariyah s pomosch'yu yacheyki bezzhidkostnogo soedineniya / P. Ya. Ti-schenko [i dr.] // Okeanologiya. 2002. T. 42, № 1. S. 32–41.
14. Handbook of methods for the analysis of the various parameters of the carbon dioxide system in sea water. Version 2 / Eds. A. G. Dickson and C. Goyet. DOE, 1994. 198 p. (OENL/CDIAC-74).
15. Atlas of Japan (East) Sea hydrographic properties in summer, 1999 / L. D. Talley [et al.] // Pro-gress in Oceanography. 2004. Vol. 61, iss. 2–4. P. 277–348. https://doi.org/10.1016/j.pocean.2004.06.011
16. Burov B. A., Lazaryuk A. Yu., Lobanov V. B. Issledovanie teplovogo potoka na granice voda-donnye osadki v Amurskom zalive Yaponskogo morya v zimniy period // Okeanolo-giya. 2014. T. 54, № 6. S. 744. EDN TCJAJZ. https://doi.org/10.7868/S0030157414060021
17. Redfield A. C., Ketchum B. H., Richards F. A. The influence of organisms on the composition of seawater // The sea: ideas and observations on progress in the study of the seas. New York, Interscience Publishers, 1963.Vol. 2. P. 26–77.
18. Ocenka potokov biogennyh veschestv na granice razdela voda – dno v melkovodnyh bas-seynah zaliva Petra Velikogo (Yaponskoe more) / P. Ya. Tischenko [i dr.] // Geohimiya. 2022. T. 67, № 9. S. 881–895. EDN ILJBJX. https://doi.org/10.31857/S0016752522090084
19. Vyshkvarcev D. I., Konovalova G. V. O mehanizmah zimnego «cveteniya» mikrovodoros-ley v vodah zaliva Petra Velikogo (Yaponskoe more) // Biologiya morya. 1979. № 1. S. 72–73.
20. Diagenez organicheskogo veschestva verhnego sloya donnyh otlozheniy zaliva Petra Veli-kogo v mestah proyavleniya gipoksii / P. Ya. Tischenko [i dr.] // Geohimiya. 2018. № 2. S. 185–196. EDN YTFMTH. https://doi.org/10.7868/S0016752518010090
21. Hidden seafloor hypoxia in coastal waters / J. P. Fredriksson [et al.] // Limnology and Ocean-ography. 2024. Vol. 69, iss. 11. P. 2489–2502. http://dx.doi.org/10.1002/lno.12607