














Sevastopol, Russian Federation
UDC 581.132
Purpose. The purpose of the study is to present an algorithm for calculating the integral phytoplankton biomass in the Black Sea euphotic layer using expeditionary data, and to perform a comparative analysis of the variability of studied characteristics obtained by means of calculations in two ways: using direct measurements of chlorophyll concentration along the horizons, and based on the parameterization re-sults. Methods and Results. The algorithm for calculating the integral biomass of phytoplankton is presented. The data on the Crimea coast waters at the 20–1500 m depths for different seasons used in the study were obtained during the cruises of R/V “Professor Vodyanitsky” in 2018–2022. The estimates resulted from parameterization and those obtained from calculations based on direct measurements of the indi-vidual input parameters at different depths are compared. The results of parameterization statistical anal-ysis show that the determination coefficients varied in the range 0.7–0.74. In the photosynthesis zone, the monthly averages of integral phytoplankton biomass (calculated from the expeditionary data) in June and October constitute 768 ± 283 and 2277 ± 726 mg C/m2, respectively. In the upper mixed layer, in June they are 556 ± 270 mg C/m2, and in October – 2023 ± 725 mg C/m2. The parameterization-derived monthly averages for the whole water area under study differ from the ones calculated using the direct measurements of input parameters at different depths by 0.9–4%. The chlorophyll a concentration pro-files for individual months in 2018–2022 are considered and mathematically described using the func-tion obtained in earlier studies. In autumn, the maximum values of chlorophyll a are observed mainly in the upper mixed layer, in summer – at the lower boundary of euphotic zone where up to 0.1% of light falling on the sea surface, penetrates. Conclusions. The above parameterization of integral phytoplankton biomass is applicable to all the sea-sons, is easy to use and agrees well with the results of calculations based on direct measurements of chlorophyll concentration at different depths. In future, the calculation algorithm is expected to become convenient for computations using satellite data.
integral biomass, phytoplankton, Black Sea, calculation algorithm, chlorophyll concentration profiles
Введение
Фитопланктон является первичным звеном в трофической цепи водных экосистем. Одним из важнейших его показателей может служить биомасса. Изменчивость биомассы фитопланктона будет отражаться на развитии всех последующих трофических уровней. На протяжении многих лет проводилось немало исследований изменения во времени и распределении в пространстве биомассы фитопланктона в море [1–12]. Биомассу микроводорослей определяют обычно путем прямых измерений, как правило, по объему клеток с дальнейшим перерасчетом ее в различных размерностях [1, 3, 4, 13, 14]. Кроме прямых методов определения разрабатываются модели, позволяющие оценивать отношение хлорофилла к органическому углероду и биомассу фитопланктона [9, 15–17]. Модели расчета биомассы фитопланктона с включением минимального числа легкодоступных для измерений входных параметров могут существенно упростить задачу вычисления этого показателя, особенно при анализе пространственных и временных изменений. Также модели расчета параметров фитопланктона необходимы для использования в исследованиях с применением спутниковых данных. Отдельный интерес представляют оценки интегральных характеристик фитопланктона в зоне фотосинтеза. Например, простые и удобные в использовании модели расчета интегральной первичной продукции разрабатывались для Черного моря [18, 19]. Вместе с тем подобных расчетных алгоритмов с легкодоступными входными параметрами для интегральной биомассы фитопланктона ранее в литературе было представлено мало. Модельные данные уступают в точности прямым измерениям, но прямые методы оценок довольно трудоемки. Расчетный способ определения интегральной биомассы фитопланктона в эвфотической зоне (ЭФЗ) и верхнем квазиоднородном слое (ВКС) позволит проводить оперативный и обширный анализ экосистемы водоема и использовать его в расчетах с данными спутниковых наблюдений.
Цель работы – представить простой алгоритм расчета интегральной биомассы в эвфотическом слое с применением экспедиционных данных и провести сравнительный анализ изменчивости исследуемых характеристик по вычислениям с помощью прямых измерений отдельных входных параметров и результатов параметризации.
Материалы и методы
Использованные в работе измерения выполнены в Центре коллективного пользования НИС «Профессор Водяницкий» во время 122-го и 124-го рейсов 7 июня – 2 июля и 3–20 октября 2022 г. в Черном море у южного и юго-восточного побережий Крыма. Общая глубина станций составляла 32–1500 м. Горизонты для отбора проб выбирали через 10–20 м от поверхностного слоя, а также с учетом границы термоклина на глубоководных станциях и придонного слоя на станциях общей глубиной ≤ 100 м.
Интенсивность падающего на поверхность моря света измеряли регистратором освещенности LI-1500 (LI-COR, США), оснащенным квантовым датчиком LI-190R (США), с 08:00 до 20:00 ежедневно, затем рассчитывали интегральное суточное значение.
Верхний квазиоднородный слой рассчитывали с использованием данных зонда IDRONAUT OCEAN SEVEN 320 Plus M как глубину, на которой относительная плотность воды на 0,07 выше, чем на поверхности [20].
Относительную прозрачность воды определяли с помощью диска Секки в дневное время суток. На основе данных глубины видимости белого диска Секки (Zs) определяли глубину ЭФЗ (Zeu = 3Zs) куда проникает до 1% света, падающего на поверхность, и kd (м–1) – показатель диффузионного ослабления света, который оценивали по формуле, полученной по данным работ
kd = 4,6/Zeu (r2 = 0,96).
Методика определения концентрации хлорофилла (далее в настоящей работе имеется в виду хлорофилл а), данные измерений, проведенных в рейсах, а также исследуемые акватории описаны в работе [21].
Для анализа профилей концентрации хлорофилла использовались данные, полученные в 2018–2022 гг. на НИС «Профессор Водяницкий» в 105, 106, 108, 110, 122, 124-м рейсах в ноябре – декабре, апреле, июле – августе, октябре, июне и октябре соответственно. Измерения концентрации проводили, как правило, через каждые 10 м до глубины 40–50 м и в отдельных случаях глубже.
Статистическая обработка данных проводилась с использованием программ Excel, SigmaPlot, Grapher, OriginLab.
Результаты
На основе разработанного нами ранее алгоритма расчета биомассы фитопланктона для поверхностного слоя Черного моря [15], скорректированного в работе [22], в настоящем исследовании проведена его модификация для оценки интегральных показателей. Вычислялись входные параметры для каждого горизонта, затем биомасса интегрировалась по глубине.
Использовалось уравнение для расчета биомассы фитопланктона Вz (мг С/м3) на глубине z (м):
Вz = Хлz/Хл:Сz, (1)
где Хлz (мг/м3) – концентрация хлорофилла на глубине z; Хл:Сz – отношение концентрации хлорофилла к органическому углероду, которое рассчитывалось на каждом горизонте:
Хл:Сz = 0,0072(Ezaph z)–0,395 (r2 = 0,78), (2)
aph z = 0,017Хлz–0,29. (3)
Параметры уравнения для Хл:С были получены на культурах водорослей в лабораторных экспериментах, описанных в работе [15], при различной освещенности. Для оценки Хл:С на разных горизонтах освещенность на каждой глубине определялась по уравнению
Ez = 0,94Е0ехр(–kdz), (4)
где Еz, Е0 (моль квантов/м2∙сут) – интенсивность фотосинтетически активной радиации (ФАР), падающей на поверхность моря и на глубину z; коэффициент 0,94 вводится для учета отражения 6% света от морской поверхности ; aph z (м2/мгХл) – показатель поглощения света пигментами водорослей, нормированный на концентрацию хлорофилла.
Для расчета интегральной биомассы фитопланктона применялись уравнения распределений необходимых показателей. Для концентрации хлорофилла использовалась кривая Гаусса, как предложено в работе [23]:
Хлz = (h/σ(2π)1/2)exp[– (z – zм)2/2σ2], (5)
h = (55,73 ± 1,40)Хл0(0,56 ± 0,008) (r2 = 0,75), (6)
zм = (11,1 ± 0,75) – (10,46 ± 0,45)ln(Хл0) (r2 = 0,61), (7)
где h – общее содержание хлорофилла (мг/м3) в слое максимума; σ (м) – показатель ширины глубинного максимума хлорофилла; zм (м) – глубина максимума хлорофилла; Хл0 (мг/м3) – концентрация хлорофилла в поверхностном слое. При оценке глубины максимума хлорофилла, согласно функции (7), вносится допущение, при котором максимум будет находиться все время на поверхности, если Хл0 > 2,89 мг/м3, однако при расчете уравнения (5) zм используется в виде функции (7). Ширина максимума σ в среднем равна 20 ± 10 м при Хл0 < 1 мг/м3 и 13 ± 8 м при Хл0 > 1мг/м3 [23]. Она рассчитывалась при значениях, равных 68% от высоты пика хлорофилла, согласно закону распределения Гаусса. Формула (5) с входными компонентами справедлива для значений концентрации хлорофилла в поверхностном слое, характерных для Черного моря. Данное распределение свойственно для теплого периода года (апрель – октябрь, иногда ноябрь); для холодного периода (декабрь – март) считается, что хлорофилл распределяется равномерно [18, 23]. Условия питания при разработке модели принимались оптимальными.
Для каждой глубины рассчитывались показатели aph z, Хл:Сz, Вz, Еz с учетом прямых измерений, полученных в рейсах. Расчет интегральной биомассы велся двумя способами: первый включал в алгоритм прямые измерения концентрации хлорофилла, второй – уравнение (5) распределения данного параметра с глубиной; остальные параметры рассчитывались идентично. Далее биомасса интегрировалась по глубинам для ЭФЗ и ВКС.
Результаты сравнения интегральной биомассы фитопланктона, полученной расчетным путем по представленному алгоритму с включением уравнения (5), и биомассы, рассчитанной по данным прямых измерений концентрации хлорофилла, показали высокую согласованность для прибрежной области вдоль Крыма и открытого прибрежья (глубины более 500 м) северной части Черного моря. Статистические характеристики сравниваемых величин для зоны фотосинтеза и верхнего квазиоднородного слоя представлены на рис. 1 и в таблице.
Статистические показатели и средние значения, полученные по алгоритму
и вычисленные с помощью прямых измерений концентрации хлорофилла
на горизонтах для интегральной биомассы фитопланктона, осредненной
по данным 122-го и 124-го рейсов НИС «Профессор Водяницкий»
Statistical indicators and average values obtained by the algorithm and calculated
using direct measurements of chlorophyll concentration at horizons for the integral phytoplankton biomass averaged based on the data collected
in the 122nd and 124th cruises of R/V “Professor Vodyanitsky”
|
Зоны / Zones |
r |
r2 |
F |
P |
<В>122 |
<В>124 |
<Вр>122 |
<Вр>124 |
|
ЭФЗ / Euphotic zone |
0,84 |
0,70 |
75,4 |
< 0,0001 |
768 ± 283 |
2277 ± 726 |
776 ± 276 |
2212 ± 759 |
|
КС / UML |
0,86 |
0,74 |
89,5 |
< 0,0001 |
556 ± 270 |
2023 ± 725 |
561 ± 240 |
1942 ± 719 |
П р и м е ч а н и е. r – коэффициент корреляции, r2 – коэффициент детерминации, F – критерий Фишера, P – уровень значимости, <В>, <Вр> (мг С/м2) – средние значения интегральной биомассы фитопланктона по данным с использованием прямых измерений и результатов параметризации соответственно.
N o t e: r is the correlation coefficient, r2 is the determination coefficient, F is the Fisher criterion, P is the importance level, <B> and <Вр> (mg C/m2) are the average values of integral phytoplankton biomass obtained using direct measurements and parameterization results, respectively.
Различия между модельными и измеренными данными профилей концентрации хлорофилла отражаются на результатах расчета биомассы двумя способами и проявляются в случаях, когда максимум концентрации хлорофилла на поверхности довольно высокий, а затем с глубиной концентрация резко снижается либо, наоборот, на поверхности находятся небольшие значения, а на глубине – максимум с высокой концентрацией хлорофилла. Существенная разница между результатами расчета биомассы двумя способами также наблюдается при наличии двухвершинных профилей концентрации хлорофилла. По сравнению с прямыми измерениями модельные расчеты могут как завышать, так и занижать биомассу фитопланктона в полтора раза и более в случаях, описанных выше. Однако количество профилей концентрации хлорофилла с такими выраженными отклонениями от описания с помощью параметризации невелико: 18% по данным двух рейсов в ЭФЗ и 15% – в ВКС. Измерения в рейсах проводились почти ежесуточно на различных станциях прибрежной и глубоководной акваторий вблизи берегов Крыма. Соответственно, при осреднении будет происходить сглаживание и приближение результатов, полученных с помощью функции распределения и по расчетам с использованием данных прямых измерений (таблица, рис. 1).
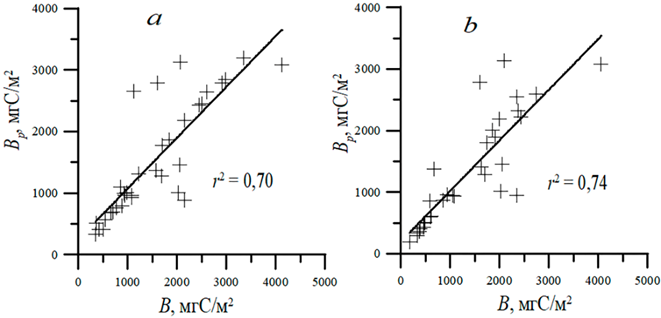
Р и с. 1. Сопоставление интегральной биомассы фитопланктона, полученной с помощью параметризации (Вр) и вычисленной по данным прямых измерений входных параметров (В) для ЭФЗ (а) и ВКС (b)
F i g. 1. Comparison of the values of integral phytoplankton biomass obtained by parameterization (Bp) and calculated using direct measurements of input parameters (B) for the euphotic zone (a) and UML (b)
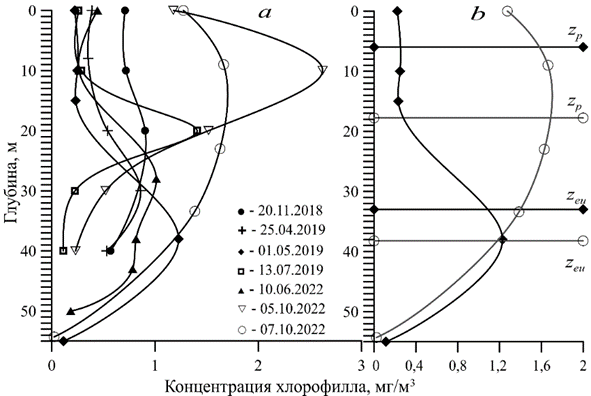
Р и с. 2. Изменение концентрации хлорофилла с глубиной по данным, полученным в рейсах НИС «Профессор Водяницкий» в 2018–2022 гг.: а – на примере отдельных дней, характерных для различных месяцев; b – на примере 07.10.2022 г. (○) и 01.05.2019 г. (♦) с границами ВКС (zp) и ЭФЗ (zeu)
F i g. 2. Change in chlorophyll concentration with depth based on the data obtained during the cruises of R/V “Professor Vodyanitsky” in 2018–2022: a – on the example of individual days typical of different months; b – on the examples of 07.10.2022 (○) and 01.05.2019 (♦) with the boundaries of UML (zp) and euphotic zone (zeu)
В работе проанализировано 88 профилей концентрации хлорофилла в различные месяцы с апреля по декабрь. Для примера на рис. 2 приведены некоторые из них. За весь рассмотренный период двухвершинные профили встречались всего 9 раз, трехвершинный – один раз. Такие профили наблюдались преимущественно в октябре (6 раз), а также в июне (3 раза) и августе (один раз). Максимум концентрации хлорофилла наблюдался в пределах зоны ВКС обычно в октябре, ноябре и декабре (рис. 2). Однако в октябре в 110-м рейсе максимум чаще был отмечен на нижней границе ВКС или ниже этой зоны. В июне и апреле максимум концентрации хлорофилла находился ниже зоны ВКС на нижней границе эвфотического слоя (рис. 2) и иногда ниже 1% освещенности от ФАР, падающей на поверхность моря. В июле – августе пик хлорофилла также наблюдался в пределах эвфотической зоны или на ее нижней границе. Уравнение (5) распределения концентрации хлорофилла [23], представленное выше, давало близкое описание профилей к данным измерений. По нашим данным с использованием всех точек в двух рейсах тестирование показало в июне r = 0,68, в октябре r = 0,64 (рис. 3). Среди всех рассмотренных профилей концентрации хлорофилла за два рейса (122-й и 124-й) (в расчетах использовалось 218 точек) коэффициент корреляции изменялся для 21 профиля в диапазоне 0,80–0,99, для пяти профилей – в диапазоне 0,60–0,8, для пяти профилей – в диапазоне 0,30–0,60 и для трех профилей он был менее 0,10.
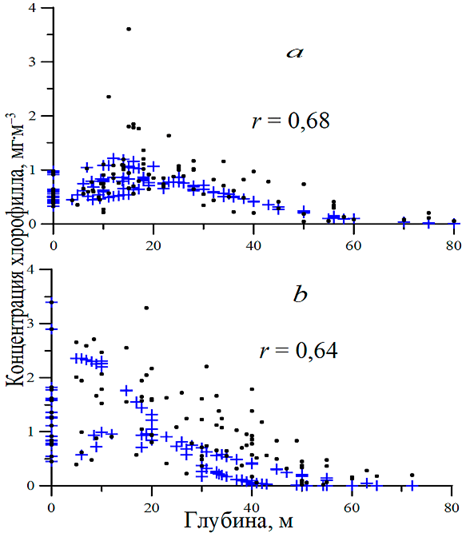
Р и с. 3. Изменение концентрации хлорофилла с глубиной в 122-м (а) и 124-м (b) рейсах НИС «Профессор Водяницкий» (данные, полученные по функции распределения (5), показаны крестиком, данные прямых измерений – кружочком)
F i g. 3. Change in chlorophyll concentration with depth in the 122nd (а) and 124th (b) cruises of R/V “Professor Vodyanitsky” (data obtained using the distribution function (5) are indicated with a cross, and direct measurement data – with a circle)
Рассматривалась связь σ с Хл0, глубиной ВКС и ЭФЗ, температурой в поверхностном слое моря, а также с E0 и kd. Наилучшая согласованность из всех рассмотренных параметров для показателя σ наблюдалась с ВКС, глубиной ЭФЗ и Хл0. Однако достоверной корреляционной связи с анализируемыми параметрами в выборке выявлено не было. Согласно результатам множественной корреляции, влияние трех указанных параметров объясняло изменчивость σ на 20%, тогда как шести параметров – на 25%. Среднее значение σ составляло 25 ± 10 м за все рассмотренные месяцы, изменяясь в диапазоне 2–48 м. В осенние месяцы ширина σ была сопоставима со значениями ВКС, летом она могла превышать их в несколько раз. По сравнению с шириной ЭФЗ параметр σ почти всегда был меньше либо соизмерим с ней. Можно предположить, что σ будет зависеть от количества питательных веществ и особенностей гидрологических условий, так как с рассмотренными шестью параметрами явной корреляционной связи не обнаружено. Указанные параметры рассматривались для оценки σ в связи с тем, что они легкодоступны для определения и расчетов с помощью спутниковых данных с целью более обширных исследований в перспективе.
Также в результате исследований профилей концентрации хлорофилла выявлено, что его максимум для весеннего (с апреля) и летнего периодов может наблюдаться примерно до проникновения 0,1% света от значений на поверхности. Соответственно, граница ЭФЗ в эти месяцы будет ниже 1% ФАР, падающей на поверхность.
Обсуждение
Данных по интегральной биомассе фитопланктона в литературе представлено довольно мало, особенно для Черного моря. Так, например, авторы работы [24] оценивают интегральную биомассу диатомовых водорослей, исходя из моделей [17, 25] и сопоставляя результаты с концентрацией хлорофилла, полученной по спутниковым данным. Упрощенный подход расчета интегральной биомассы использовался ранее в работе [26] при анализе экспедиционных исследований, которые проводились зимой и весной. Использованные в указанных выше работах расчетные способы не учитывали распределения с глубиной некоторых важных характеристик фитопланктона и оптических показателей при оценке его биомассы. Расчеты проводились для разных районов, и сопоставить между собой полученные результаты было проблематично.
В работе [23] представлена функция распределения биомассы фитопланктона. Для нее требуются измерения по горизонтам концентрации нитратов и температуры воды, которые не всегда можно провести. Другая функция из этой работы, для которой требуется определение оптической глубины, дает грубые оценки по сравнению с данными прямых измерений интегральной биомассы, на что указывается самими авторами. Для примера нами было проведено сравнение расчетных данных интегральной биомассы фитопланктона по функции из указанной работы, включающей в качестве входного параметра значения оптической глубины и интегральной биомассы, полученной по нашему алгоритму с включением в расчет прямых измерений концентрации хлорофилла по горизонтам. Было проанализировано 34 профиля биомассы, полученных по результатам 122-го и 124-го рейсов НИС «Профессор Водяницкий» в июне и октябре. Их сопоставление показало согласованность двух способов расчета для июня (r2 = 0,63) и октября (r2 = 0,19). В результате установлено, что по сравнению с нашими расчетами значения, полученные по функции биомассы из работы [23] в октябре, часто были выше приблизительно в 2 раза.
Прямые определения биомассы фитопланктона, особенно интегральной, трудоемки и проводятся крайне редко. Последние 10–15 лет в Черном море они не проводились. Поэтому получить данные прямых измерений интегральной биомассы или сопоставить их с результатами расчетов не представляется возможным в связи с отсутствием таких измерений. Согласованность измеренных значений биомассы фитопланктона и расчетных, полученных по используемому нами алгоритму, для поверхностного слоя описывалась ранее в работах [15, 22, 27]. Также проведено сопоставление функции распределения (5) концентрации хлорофилла с данными прямых измерений на горизонтах в настоящей работе и в работе [23]. Исходя из указанной согласованности, мы предполагаем адекватность расчетных значений на горизонтах с учетом разной освещенности.
Наши исследования являются продолжением работ [15, 23]. Представленный здесь алгоритм расчета интегральной биомассы, основанный на лабораторных и экспедиционных исследованиях, простой в применении и показывает хорошую сопоставимость с данными расчетов, включающими прямые измерения концентрации хлорофилла на разных горизонтах (таблица). В алгоритме учитываются распределения по глубине таких важных входных параметров, как Хл, aph, Хл:С и изменение освещенности с глубиной. Используемый нами алгоритм для поверхностного слоя разрабатывался на основе 10 видов водорослей, встречающихся в Черном море (Nitschia sp., Pseudonitschia delicatissima, Skeletonema costatum, Talasiossira parva, Coscinodiscus granii, Phaeodactilum tricornutum, Prorocentrum micans, Isochrysis galbana, Dunaliella tertiolecta, Glenodinium foliaceum), в том числе из доминирующих таксономических групп (диатомовые и динофлагелляты), которые были доступны для экспериментальных работ. Для указанных видов были получены описанные выше зависимости физиологических и структурных показателей микроводорослей и средние коэффициенты для них (уравнения (1) – (7)). Условия питания в данной модели принимаются оптимальными при разной освещенности.
В нашем алгоритме мы опираемся на измерения концентрации хлорофилла и полученную ранее зависимость для значений его среднего удельного содержания в клетке для разных видов водорослей отдельных групп. Оценка Хл:С в модели из работы [15] осуществлялась с учетом поглощения света микроводорослями, что позволяет примерно оценивать общую биомассу фитопланктона, несмотря на отсутствие иных массовых видов в параметризации.
К слабой стороне нашего алгоритма можно отнести отсутствие учета кокколитофорид, которые доминируют в начале летнего периода [11, 28–30]. Поэтому для периода их «цветения» значения биомассы фитопланктона при расчете могут быть занижены. По результатам наших исследований, представленных ранее в работе [21], в июне доминировали кокколитофориды на фоне остальных групп микроводорослей. Данные интегральной биомассы, рассчитанные двумя способами, показали высокую согласованность, при этом значения биомассы фитопланктона не были низкими. Это можно объяснить относительно близкими средними значениями удельного содержания хлорофилла в клетке у учтенных нами диатомовых водорослей и кокколитофорид. Алгоритм дает общие оценки биомассы фитопланктона без разделения на группы и виды микроводорослей. Поэтому результаты стоит анализировать, принимая во внимание слабые стороны и допущения данной модели.
Анализируя профили концентрации хлорофилла, можно отметить, что в большинстве случаев образование его максимума в зоне ВКС происходит осенью. Это может быть связано с усилением перемешивания водных масс, поступлением биогенных веществ в верхние слои воды и расширением зоны ВКС. В то же время в поверхностном слое остается растворенная органика, возникшая в результате жизнедеятельности морских организмов. Уменьшение солнечной радиации также способствует тому, что зона фотосинтеза поднимается выше и виды водорослей, которые испытывали фотоингибирование в летний период в зоне ВКС, могут развиваться. Эти же причины могут обусловливать и образование двух пиков в профилях хлорофилла в этот период. Образование максимумов хлорофилла в апреле и в летний период за пределами ВКС, на нижней границе ЭФЗ, можно связать преимущественно с высоким уровнем освещенности, выраженной температурной стратификацией в столбе воды и узкой зоной ВКС. В июне, например, встречались пики профилей хлорофилла ниже 1% освещенности, что свидетельствует о расширении ЭФЗ вследствие высокого уровня ФАР, падающей на поверхность моря.
Проблема математического описания профилей концентрации хлорофилла остается до конца не решенной в случаях, когда они имеют два, иногда даже три максимума в столбе воды либо нетипичную одновершинную форму. Задача упрощается тем, что сложные профили встречаются не часто и в процессе осреднения различия сглаживаются. Это же происходит и при пересчете других параметров фитопланктона.
При построении моделей для Черного моря с учетом множества факторов, таких как гидрологические условия и включение питательных веществ (соединений азота, фосфора и кремния), необходимы контактные измерения вышеперечисленных параметров. В различные годы проводились исследования и сбор данных с выявлением взаимосвязи между такими параметрами, например, доминирующими видами и питательными веществами с учетом ветровой активности [12], а также взаимосвязи речного стока со структурой фитопланктонного сообщества [31]. Учет всех этих факторов в перспективе приведет к разработке сложных моделей. Для оценки интегральных величин по спутниковым данным в настоящий момент и ближайшее время будут доступны не все указанные входные параметры. Существуют сложные глобальные модели морских экосистем, включающие большое количество входных параметров [17, 32–34], однако они также не описывают все случаи, имеют свои допущения, ограничения и погрешности.
Наша задача на данный момент сводится к созданию простого алгоритма для оценки общей биомассы фитопланктона и ее интегральных значений в столбе воды с учетом средних соотношений удельного хлорофилла в клетке некоторых доминирующих групп микроводорослей (уравнение (2)). Предложенный алгоритм применим для всех сезонов года с учетом распределения концентрации хлорофилла (уравнение (5)) с апреля по ноябрь и его равномерного распределения с декабря по март [23]. Результаты анализировались на примере двух сезонов. Так, среднемесячные значения интегральной биомассы фитопланктона, полученные по нашей параметризации и по расчетам с помощью измеренных входных параметров на разных глубинах, отличаются на 0,9–4% (таблица). При ограниченном поступлении отдельных питательных веществ возможны отклонения в соотношении Хл:С. В реальных условиях также происходит смена видового состава. Подобные факторы предложенная нами параметризация не учитывает. В алгоритме возможны лишь оценки изменчивости общей биомассы фитопланктона по средним характеристикам, описанным выше (уравнения (1) – (7)). Для оценок интегральной биомассы предложенная параметризация может быть применима. Алгоритм расчета в перспективе будет удобен для вычислений с использованием спутниковых данных при учете гидрооптических характеристик для Черного моря [35].
Заключение
По представленному в работе алгоритму проведены расчеты интегральной биомассы фитопланктона для прибрежной зоны и открытого прибрежья (глубины свыше 500 м) северной части Черного моря. Хорошо согласуются результаты расчета интегральной биомассы, полученные двумя способами: с использованием данных прямых измерений входных параметров на разных глубинах в экспедиционных исследованиях и с помощью уравнений распределения входных параметров. Согласно проведенной статистической обработке, для данных двух рейсов коэффициенты детерминации равны 0,7 и 0,74. Среднемесячные значения интегральной биомассы фитопланктона и стандартные отклонения, рассчитанные по результатам экспедиционных исследований в зоне фотосинтеза для июня и октября, равны 768 ± 283 и 2277 ± 726 мг С/м2 соответственно, а в верхнем квазиоднородном слое они составляют 556 ± 270 мг С/м2 в июне и 2023 ± 725 мг С/м2 в октябре. Осредненные месячные значения по расчетам двумя способами отличались на 0,9–4%.
Приведенный алгоритм, включающий уравнения распределения всех входных параметров, прост в использовании и будет удобен при работе со спутниковыми данными. Из анализа профилей концентрации хлорофилла выявлено, что его пик в осенний период наблюдался обычно в зоне ВКС, тогда как в апреле – июле – на нижней границе эвфотического слоя, куда проникает до 0,1% от ФАР, падающей на поверхность моря. В результате множественного корреляционного анализа не было выявлено достоверной корреляционной связи между шириной глубинного максимума концентрации хлорофилла в слое воды и шестью параметрами – концентрацией хлорофилла на поверхности, глубиной ВКС и эвфотического слоя, температурой и освещенностью на поверхности моря, коэффициентом диффузного ослабления света. Получено, что влияние указанных параметров на изменчивость ширины максимума хлорофилла составляло 25%.
1. Selifonova Zh. P., Yasakova O. N. Fitoplankton akvatoriy portovyh gorodov severo-vostochnogo shel'fa Chernogo morya // Mors'kiy ekologichniy zhurnal. 2012. T. 11, № 4. S. 67–77. EDN TMKHFV.
2. Vliyanie kompleksa faktorov sredy na biomassu fitoplanktona i zooplanktona v Cher-nom more v vesenniy period / S. B. Krasheninnikova [i dr.] // Ekologiya i stroitel'stvo. 2019. T. 4. S. 14–21. EDN RAWNQO. https://doi.org/10.35688/2413-8452-2019-04-002
3. Stel'mah L. V., Mansurova I. M. Mnogoletnyaya dinamika fitoplanktona i koncentracii hlorofilla a v poverhnostnom sloe pribrezhnyh vod Chernogo morya (rayon Sevastopo-lya) // Voprosy sovremennoy al'gologii. 2020. № 1. S. 66–81. EDN NBMFBU. https://doi.org/https://doi.org/10.33624/2311-0147-2020-1(22)-66-81
4. Stel'mah L. V. Vliyanie adaptacii fitoplanktona na raspredelenie ego biomassy i kon-centracii hlorofilla a v poverhnostnom sloe Chernogo morya // Sistemy kontrolya okru-zhayuschey sredy. 2019. Vyp. 1. S. 106–114. https://doi.org/10.33075/2220-5861-2019-1-106-114
5. Morozova-Vodyanickaya N. V. Fitoplankton Chernogo morya. Chast' I. Fitoplankton v rayone Sevastopolya i obschiy obzor fitoplanktona Chernogo morya // Trudy Sevasto-pol'skoy biologicheskoy stancii. Moskva – Leningrad : Izd-vo AN SSSR, 1948. T. VI. S. 39–172.
6. Morozova-Vodyanickaya N. V. Fitoplankton Chernogo morya. Chast' II // Trudy Seva-stopol'skoy biologicheskoy stancii. Moskva – Leningrad : Izd-vo AN SSSR, 1954. T. VIII. S. 11–99.
7. Basin-scale variability of phytoplankton biomass, production and growth in the Atlantic Ocean / E. Marañón [et al.] // Deep Sea Research Part I: Oceanographic Research Papers. 2000. Vol. 47, iss. 5. P. 825–857. https://doi.org/10.1016/S0967-0637(99)00087-4
8. Vertical distribution of phytoplankton biomass, production and growth in the Atlantic subtropical gyres / V. Pérez [et al.] // Deep Sea Research Part I: Oceanographic Research Pa-pers. 2006. Vol. 53, iss. 10. P. 1616–1634. https://doi.org/10.1016/j.dsr.2006.07.008
9. Carbon-based ocean productivity and phytoplankton physiology from space / M. J. Behrenfeld [et al.] // Global Biogeochemical Cycles. 2005. Vol. 19, iss. 1. GB1006. https://doi.org/10.1029/2004GB002299
10. Stelmakh L. V. Spatial and Temporal Variability of Carbon to Chlorophyll a Ratio in Phyto-plankton of the Surface Layer in Shallow Water Areas of the Black Sea (Crimea) // In-ternational Journal on Algae. 2015. Vol. 17, iss. 4. P. 385–396. https://doi.org/10.1615/InterJAlgae.v17.i4.60
11. Alternation of diatoms and coccolithophores in the north-eastern Black Sea: a response to nutri-ent changes/ A. S. Mikaelyan [et al.] // Hydrobiologia. 2015. Vol. 755. R. 89–105. https://doi.org/10.1007/s10750-015-2219-z
12. Drivers of phytoplankton blooms in the northeastern Black Sea / V. A. Silkin [et al.] // Marine Pollution Bulletin. 2019. Vol. 138. P. 274–284. https://doi.org/10.1016/j.marpolbul.2018.11.042
13. Struktura fitoplanktona Belogo morya posle letnego cveteniya: prostranstvennaya neod-norodnost' v zavisimosti ot gidrofizicheskih usloviy / L. S. Zhitina [i dr.] // Sibir-skiy ekologicheskiy zhurnal. 2016. T. 23, № 6. S. 888–899. EDN XCRXPT. https://doi. org/10.15372/SEJ20160608
14. Menden-Deuer S., Lessard E. J. Carbon to volume relationships for dinoflagellates, diatoms, and other protist plankton // Limnology and Oceanography. 2000. Vol. 45, iss. 3. P. 569–579. https://doi.org/10.4319/lo.2000.45.3.0569
15. Finenko Z. Z., Kovaleva I. V., Suslin V. V. Novyy podhod k ocenke biomassy fitoplank-tona i ee variabel'nosti v poverhnostnom sloe Chernogo morya po sputnikovym dannym // Uspehi sovremennoy biologii. 2018. T. 138, № 3. S. 294–307. EDN XUKMZV. https://doi.org/10.7868/S0042132418030079
16. Abakumov A. I., Pak S. Ya. Modelirovanie processa fotosinteza i ocenka dinamiki biomassy fitoplanktona na osnove modeli Drupa // Matematicheskaya biologiya i bioin-formatika. 2021. T. 16, № 2. S. 380–393. EDN MVYEZI. https://doi. org/10.17537/2021.16.380
17. Oguz T., Ducklow H. W., Malanotte-Rizzoli P. Modeling distinct vertical biogeochemical struc-ture of the Black Sea: Dynamical coupling of the oxic, suboxic, and anoxic layers // Global Bio-geochemical Cycles. 2000. Vol. 14, iss. 4. P. 1331–1352. https://doi.org/10.1029/1999GB001253
18. Finenko Z. Z., Suslin V. V., Churilova T. Ya. Regional'naya model' dlya rascheta pervichnoy produkcii Chernogo morya s ispol'zovaniem dannyh sputnikovogo skanera cveta SeaWiFS // Mors'kiy ekologichniy zhurnal. 2009. T. 8, № 1. S. 81–106. EDN TNCEMJ.
19. Kovaleva I. V., Suslin V. V. Integral'naya pervichnaya produkciya v glubokovodnyh rayo-nah Chernogo morya v 1998–2015 godah // Morskoy gidrofizicheskiy zhurnal. 2022. T. 38, № 4. S. 432–445. EDN MYVMZX. https://doi.org/10.22449/0233-7584-2022-4-432-445
20. Izmenchivost' tolschiny peremeshannogo sloya v Chernom more i ee svyaz' s dinamikoy vod i atmosfernym vozdeystviem / A. A. Kubryakov [i dr.] // Morskoy gidrofizicheskiy zhur-nal. 2019. T. 35, № 5. S. 449–468. EDN AIEJPU. https://doi.org/10.22449/0233-7584-2019-5-449-468
21. Structural and functional parameters of the Black Sea phytoplankton during the summer bloom of the coccolithophore Emiliania huxleyi / L. V. Stelmakh [et al.] // Regional Studies in Marine Science. 2024. Vol. 76. 103594. https://doi.org/10.1016/j.rsma.2024.103594
22. Kovaleva I. V., Suslin V. V. Sezonnaya izmenchivost' biomassy i udel'noy skorosti rosta fitoplanktona v 2016–2020 godah v glubokovodnoy zone Chernogo morya // Sovremennye problemy distancionnogo zondirovaniya Zemli iz kosmosa. 2023. T. 20, № 4. S. 250–262. EDN VXDPLZ. https://doi.org/10.21046/2070-7401-2023-20-4-250-262
23. Finenko Z. Z., Churilova T. Ya., Li R. I. Vertikal'noe raspredelenie hlorofilla i fluo-rescencii v Chernom more // Mors'kiy ekologichniy zhurnal. 2005. T. 4, № 1. S. 15–46. EDN VKGJUN.
24. Kubryakova E. A., Kubryakov A. A., Stanichnyy S. V. Vliyanie zimnego vyholazhivaniya na vertikal'noe vovlechenie vod i intensivnost' cveteniya fitoplanktona v Chernom more // Morskoy gidrofizicheskiy zhurnal. 2018. T. 34, № 3. S. 206–222. EDN VLPZDY. https://doi.org/10.22449/0233-7584-2018-3-206-222
25. Mellor G. L. One-Dimensional, Ocean Surface Layer Modeling: A Problem and a Solution // Journal of Physical Oceanography. 2001. Vol. 31, iss. 3. P. 790–809. https://doi.org/10.1175/1520-0485(2001)031<0790:ODOSLM>2.0.CO;2
26. Razvitie fitoplanktona v zimne-vesenniy period v pribrezhnyh vodah Kryma / Z. Z. Finenko [i dr.] // Morskoy biologicheskiy zhurnal. 2021. T. 6, № 1. S. 102–114. EDN BLFXXH. https://doi.org/10.21072/mbj.2021.06.1.08
27. Finenko Z. Z., Mansurova I. M., Suslin V. V. Temporal Dynamics of Phytoplankton Biomass in the Surface Layer of the Black Sea According to Satellite Observations // Oceanology. 2022. Vol. 62, iss. 3. P. 358–368. https://doi.org/10.1134/S0001437022030043
28. Pautova L. A., Mikaelyan A. S., Silkin V. A. Structure of plankton phytocoenoses in the shelf waters of the northeastern Black Sea during the Emiliania huxleyi bloom in 2002–2005 // Oceanology. 2007. Vol. 47, iss. 3. P. 377–385. https://doi.org/10.1134/S0001437007030101
29. Mikaelyan A. S., Silkin V. A., Pautova L. A. Coccolithophorids in the Black Sea: Their interan-nual and long-term changes // Oceanology. 2011. Vol. 51, iss. 1. R. 39–48. https://doi.org/10.1134/S0001437011010127
30. Environmental control on phytoplankton community structure in the NE Black Sea / V. A. Silkin [et al.] // Journal of Experimental Marine Biology and Ecology. 2014. Vol. 461. P. 267–274. https://doi.org/10.1016/j.jembe.2014.08.009
31. Response of the Coastal Phytoplankton Community to the Runoff from Small Rivers in the Northeastern Black Sea / V. M. Sergeeva [et al.] // Diversity. 2023. Vol. 15, iss. 7. 857. https://doi.org/10.3390/d15070857
32. Vichi M., Pinardi N., Masina S. A generalized model of pelagic biogeochemistry for the global ocean ecosystem. Part I: Theory // Journal of Marin Systems. 2007. Vol. 64, iss. 1–4. P. 89–109. https://doi.org/10.1016/j.jmarsys.2006.03.006
33. Dorofeev V. L., Korotaev G. K., Suhih L. I. Cistema diagnoza-prognoza sostoyaniya eko-sistemy Chernogo morya // Problemy ekologicheskogo monitoringa i modelirovaniya eko-sistem. 2017. T. 28, № 2. S. 71–85. EDN ZGSVTP. https://doi.org/10.21513/0207-2564-2017-2-71-85
34. Belyaev V. I., Konduforova N. V. Matematicheskoe modelirovanie ekologicheskih sistem shel'fa. Kiev : Naukova dumka, 1990. 239 s.
35. Suslin V. V., Churilova T. Ya., Pryahina S. F. Regional'naya metodika vosstanovleniya per-vichnyh gidroopticheskih harakteristik Chernogo morya po dannym cvetovogo skanera SeaWiFS // Ekologicheskaya bezopasnost' pribrezhnoy i shel'fovoy zon i kompleksnoe is-pol'zovanie resursov shel'fa. 2012. Vyp. 26, t. 2. S. 204–223. EDN VWSEIV.